生物竞赛补充
总纲
写的时候要过脑子而不是过书。不管如何先看完书再打。看书的时候要动脑子。机制类的先简化后填充,死记硬背类的先跳过后来再来打。
生物化学
核酸的理化性质和研究方法
- 核酸变性因素:高温、碱性pH,低离子强度,有机溶剂(甲酰胺、甲醛、甲醇、乙醇、尿素)
氨基酸
- 只要含有游离氨基和羧基的物质都可以发生茚三酮反应,如PS。环肽不能。
- 氨基酸和亚硝酸或甲醛的反应和用途。氨基酸的氨基可以和甲醛在有硼氢化钠类还原剂存在的条件下发生还原胺化反应,在氨基上上一个甲基。这是在氨基上定量甲基化的重要方法。和亚硝酸反应生成氮气,氮气中的氮素一半来自于氨基酸,凯氏定氮法。
- 和氨基酸侧链集团有关的反应和用途。
- Arg的胍基——坂口试剂【alpha萘酚、次溴酸钠】——桃红色。【关于alpha-萘酚:其不仅参与了坂口试剂用于测定精氨酸;还参与了:1.Molisch反应用于检测碳水化合物的存在;2.Rapid furfural test用于检测果糖的存在;3.VP测试表明葡萄糖分解为3-羟基丁酮。】
- Met的-S-CH3——H2O2——过氧化物
- Cys的-SH——碘代乙酸、过甲酸——乙酸巯基、磺酸基
- His的咪唑基——Pauly试剂【重氮苯磺酸】——樱红色物质
- Tyr的酚羟基——Pauly试剂——桔黄色物质【HPY】
- Tyr的酚羟基——Folin试剂——蓝色物质
- Trp的吲哚基——对二甲基氨基苯甲醛——蓝色物质
- R基具有手性C的氨基酸是Ile和Thr。
代谢总论
- 代谢组 又被称为 小分子清单SMI,并不包括生物大分子。代谢组代表了细胞的当前状态。分离代谢小分子的方法有毛细管电泳CE、气相色谱GC 和 高效液相层析。鉴定代谢小分子的方法有质谱 和 核磁共振。
- 磷酸精氨酸 和 磷酸肌酸 统称为磷酸原。某些微生物使用聚偏磷酸。
生物氧化
-
黄素通常作为辅基。因其和泛醌不同,其半醌中间物反应性极强,必须两个电子同时传递。
-
泛醌也存在于其他的生物膜上作为抗氧化剂。
-
铁硫蛋白含有非血红素铁 和 对酸不稳定的无机S。有3类:Fe,Fe2S2,Fe4S4。可以作为顺乌头酸酶、固氮酶的 辅因子。
-
复合体 III 通常形成二聚体。复合体IV 中的 血红素a3 和 CuB形成 双核中心,催化氧气的还原反应,氧气的部分还原产物和双核中心紧密结合知道水分子的形成。
-
通过蓝色非变性聚丙烯酰胺凝胶电泳BN-PAGE,分离出呼吸体。蓝色是因为加入了考马斯亮蓝。
-
厌氧生物呼吸链的末端电子受体
大肠杆菌 除硫单胞菌 醋酸杆菌 甲烷菌 脱氮副球菌 延胡索酸 S CO2 CO2 NO3- 琥珀酸 H2S 醋酸 CH4 NO2- -
复合体III 产生质子梯度的机制是 Q循环。QH2的两个电子,一个给 Cyt c,另一个给 Q 或者 Q-,并同时泵出两个质子。复合体 I 产生质子梯度的机制是分子蒸汽机。
-
寡霉素 和 DCCD 可以直接作用于 Fo 抑制ATP的合成。质子通过 Fo 主要是由 a亚基 和 C单位 组成的。有两个氨基酸至关重要——第一个是 c亚基 上的酸性氨基酸,第二个是 a亚基 上的 Arg210。a 亚基具有两个半通道。DCCD 可以共价修饰酸性氨基酸的羧基。
-
缬氨霉素 作为钾离子载体破坏电势能。IF1 可以抑制 F1Fo-ATP合酶 水解ATP。DNP, FCCP, 双香豆素可以作为解偶联剂。甲状腺激素和阿司匹林一定程度上也可以。
-
游离脂肪酸可以和CoQH2,嘌呤核苷酸一起激活UCP产热素。植物中的PUMP可以防止产生过多的ROS。
糖酵解
-
2DG是磷酸己糖异构酶的抑制剂,其催化机制涉及烯二醇中间体,而2DG不能。
-
PFK-1是最重要的限速步骤。在植物、某些细菌、古菌和少数原生动物内,具有一种PPi-PK用PPi代替ATP。此外还有部分古菌用ADP 代替ATP。
-
醛缩酶有两类:一种存在于真菌和细菌,依赖Zn++金属催化。另一类存在于其他真核生物,共价催化【涉及希夫碱中间体】。
-
TIM 是一个近乎完美的酶。
-
GADPH 是一个巯基酶,形成共价的半缩硫醛中间体,被有机汞和碘代乙酸抑制。具有负协同性,但不是别构酶,具有4个亚基。可以作为抗氧化开关进入PPP。砷酸作用类似解偶联剂。
-
磷酸甘油酸变位酶有两类,一类直接分子内转移,另一类需要2,3-BPG 作为辅因子。催化2,3-BPG 产生的酶是 1,3-BPG变位酶,其是双功能酶,还催化2,3-BPG 的水解。BPG 支路少经过一步底物水平磷酸化。但在不是RBC的一般细胞内2,3-BPG的含量低。
-
氟化物可以和烯醇化酶的 Mg++ 和磷酸基团形成络合物。
-
注意3-磷酸甘油穿梭系统 的 3-磷酸甘油脱氢酶位于 细胞质基质和mt内膜上,并产生FADH2。而苹果酸-天冬氨酸穿梭系统 的 苹果酸脱氢酶和谷草转氨酶 位于 细胞质基质和mt基质内,并产生 NADH。
-
丙酮酸 被 丙酮酸转运蛋白 和 H+ 一起进入 mt。
-
丙酮酸脱氢酶系
E1 丙酮酸脱氢酶 E2 二氢硫辛酸转乙酰酶 E3 二氢硫辛酸脱氢酶 TPP/Mg++ 硫辛酸、CoA FAD,NAD+ 丙酮酸氧化脱羧 将乙酰基转移到 CoA 氧化型硫辛酰胺的再生 -
砒霜 中 的三氧化二砷和亚砷酸 可以和还原型的硫辛酰胺 共价结合,从而抑制丙酮酸脱氢酶系 和 alpha-酮戊二酸脱氢酶系。
-
果糖经过 果糖激酶 和 1-磷酸果糖醛缩酶 裂解成DHAP 和 D-甘油醛,后者需要经过丙糖激酶形成 3-PGAld。甘露糖经过己糖激酶异构即可。甘油需要先后经过甘油激酶 和 3-磷酸甘油脱氢酶转化成 DHAP。注意有两种 3-磷酸甘油脱氢酶,这里的是产生 NADH 的类型。
-
半乳糖进入糖酵解的途径 是 Leloir 途径。参与转变的酶有 半乳糖激酶、1-磷酸半乳糖尿苷转移酶、UDP-Gal-4差向异构酶、磷酸葡糖变位酶。缺陷导致 半乳糖血症,积累有毒浓度的1-磷酸半乳糖导致。
-
视网膜和睾丸将会以糖酵解作为合成ATP的主要途径。
-
古菌细胞中 糖酵解 的变化:GK 和 PFK-1 使用ADP而不是ATP;GAPDH 使用 Fd 而不是 NADH;不经过 1,3-BPG 降低了 ATP 产量,为不可逆反应。其主要调节位点位于 GAPDH 而不是 GK 和 PFK-1.
-
糖酵解酶 的 兼职功能。如TIM作为神经白介素、自分泌;GAPDH 在细胞核作为 尿嘧啶-DNA糖苷酶;烯醇化酶参与 RNA 降解和 线粒体tRNA 的运输;醛缩酶 可以联合AMPK、V型质子泵、axin等蛋白【猴辅导出过题】。
-
葡萄糖可得性。GLUT1 组成性表达 Km 低。GLUT2 表达位于肝细胞和胰岛beta细胞 Km 高;肌细胞和脂肪细胞响应胰岛素具有GLUT4。
-
己糖激酶被 G6P 抑制,葡糖激酶被 胰岛素 和 GKRP 控制,主要合成糖原,故不受到 G6P 抑制。PFK-1 别构抑制有 ATP、柠檬酸、H+【糖“酵”解】,别构激活有 AMP,ADP,F-2.6-BP。催化 F-2.6-BP 产生的酶 PFK-2 是双功能酶,同时其C端具有 F-2,6-BPase 活性。不同PFK-2调控不同。肝细胞PFK-2酶活性主要由 胰高血糖素和胰岛素下游的 磷酸化控制。心肌细胞 PFK-2被AMPK 磷酸化响应 NE和AMP。骨骼肌细胞 PFK-2 被G6P 激活。丙酮酸激酶可以被 别构调控 和 PKA磷酸化调控;ATP,Ala别构抑制,F-1,6-BP别构激活。
-
注意 ATP 既是 PFK-1 的底物又是其别构抑制剂。其曲线呈现单峰。
三羧酸循环
- 柠檬酸合酶 催化 克莱森酯缩合反应。诱导契合模型。
- 顺乌头酸酶 含有 铁硫蛋白,但其直接参与催化反应而不是电子传递。细胞质基质也具有顺乌头酸酶用于感受Fe+++。因为假手性,异柠檬酸的羟基只和来自OAA的C相连。最终将柠檬酸的 3级羟基 转变为 容易氧化的 2级羟基。
- 异柠檬酸脱氢酶 有 IDH1,2,3。TCA循环主要是 IDH3.IDH1存在于细胞质基质和过氧化物酶体。IDH1,2参与谷氨酰胺的代谢。
- 亚砷酸也可以抑制alpha酮戊二酸脱氢酶系。其 E3 和丙酮酸脱氢酶系完全相同。
- 动物组织的 琥珀酰CoA合成酶 主要产生 GTP,而植物和微生物组织的 主要产生 ATP。
- 丙二酸抑制琥珀酸脱氢酶。
- 柠檬酸在细胞质基质可以作为别构效应物,抑制PFK-1,激活FBPase1 和 ACC。
- 一些厌氧古菌和细菌可以利用 rTCA 同化 CO2,注意其 ATP柠檬酸裂合酶,延胡索酸还原酶,alpha 酮戊二酸合酶【Fdred 而不是 NADH】。
- 乙醛酸循环 具有 苹果酸合酶 和 异柠檬酸裂合酶,只有 NADH 产生。植物细胞中的异柠檬酸脱氢酶具有磷酸化调控。
- TCA 循环的回补。OAA回补包括 PEP羧化酶【细菌、酵母、高等植物】、丙酮酸羧化酶【动物的肝和肾】、PEP羧激酶【动物心肌和骨骼肌】。alpha酮戊二酸回补包括谷丙转氨酶、谷氨酸脱氢酶。琥珀酰-CoA回补如Ile、Val、Met、Thr和奇数脂肪酸。苹果酸回补需要 苹果酸酶,并需要NADPH。
- Ca++是肌肉收缩的信号,可以作为 IDH、alpha酮戊二酸脱氢酶系、丙酮酸脱氢酶激酶 的别构激活剂。alpha酮戊二酸脱氢酶系 可以检测细胞内的自由基水平,通过谷胱甘肽化可逆抑制。双氯乙酸可以抑制丙酮酸脱氢酶激酶。
磷酸戊糖途径
- 许多癌细胞G6PDH 乙酰化修饰活性增加。转酮酶 需要 TPP。而转醛酶类似醛缩酶,涉及希夫碱中间体。
- 植物的 PPP 发生在 质体。
- ED途径。比糖酵解的产生更加古老。一个葡萄糖产生一个ATP【比糖酵解少一个】,一个NADH和【GAPDH】一个NADPH【类似PPP,在G6P脱氢产生6P葡糖酸】。经过2-酮-3-脱氧-6-磷酸葡糖酸,先产生一分子丙酮酸和一分子PGAld【后来也变成丙酮酸】。
- PPP和蚕豆病。G6PD缺乏导致蚕豆病,PPP是红细胞产生NADPH的唯一途径,因此难以维持GSH的再生。蚕豆中富含氧化剂,产生强自由基。导致红细胞遭受氧化胁迫而被破坏。然而G6PD缺乏症患者对疟疾有一定抵抗能力。
- 细胞色素P450单加氧酶解毒系统 需要 NADPH。巨噬细胞具有 NADPH氧化酶 产生超氧阴离子杀死入侵的微生物。
糖异生
- 丙酮酸羧化酶 位于 线粒体基质,需要生物素, 包括生物素羧基载体蛋白、生物素羧化酶、羧基转移酶。PEPCK 在人体内 线粒体基质和细胞质基质 都有,不同生物定位不同。
- G6P磷酸酶 位于 ER。
- 乙醇在体内可以抑制糖异生。
脂代谢
- FFA和血浆中的清蛋白结合。细胞内的长链FFA大多数和特定的蛋白结合,真正游离的很少。
- 脂肪的合成位于sER。甘油磷脂合成在sER细胞质基质一侧。鞘脂合成在sER和高尔基体,水解在溶酶体。
- TZD【噻唑烷二酮类】是治疗II型糖尿病的药物。通过激活甘油激酶,刺激脂肪合成,降低血液中的FFA浓度。
- 4种磷脂酶:A1【哺乳动物】,A2【蛇毒和蜂毒;作用于PI释放花生四烯酸;胰液被胰蛋白酶和胆盐激活;糖皮质激素抑制】,C【信号转导;杆菌α-毒素和肝细胞溶酶体】,D【植物】。
- 鞘磷脂水解在溶酶体。鞘脂酶【尼曼-皮克病】,神经酰胺酶。
- 甘油磷脂合成在sER细胞质基质一侧【磷脂爬行酶可以转移到内侧】。PI和心磷脂活化磷脂酸,消耗两分子高能磷酸键。PC,PE,PS活化头部基团,需要ATP和CTP,消耗三分子高能磷酸键。PE,PC,PS之间可以补救合成。
- 古菌具有醚键的缩醛磷脂。具有异戊二烯分支。甘油磷酸骨架为S型【细菌和真核生物为R型】。
- 鞘磷脂合成在sER和高尔基体。原料是软脂酰CoA和Ser。需要NADPH合成二氢鞘氨醇,产生神经酰胺产生FADH2。
- 鞘糖脂水解在溶酶体。鞘磷脂酶缺陷,尼曼皮克病。β半乳糖苷酶缺乏,Krabbe病。葡糖脑苷脂酶缺乏,戈谢病。【红细胞中。α半乳糖苷酶缺乏,Fabry病。β-己糖酰胺酶缺乏,Sandhoff病。】【CNS中。己糖酰胺酶缺乏,泰萨克病。】其中泰萨克病和戈谢病是常见的AR。
- 鞘糖脂合成需要活化单糖。大多为UDP-单糖。少数为CMP-单糖【如乙酰神经氨酸CMP-NeuAc】。
脂肪酸代谢
- 人类第一次使用示踪研究物质代谢:苯环标记,β氧化。
- 活化长链脂肪酸的脂酰CoA合成酶位于线粒体外膜,细胞质基质侧;短链的位于线粒体内膜,线粒体基质侧。
- 肉碱是羟基三甲胺丁酸。合成前体是Lys和Met。需要铁和维生素C,PP,B6。作为脂酰基的中间受体。L型有活性。CPT-I搭载,位于线粒体外膜;CPT-II卸下,位于线粒体内膜。
- β氧化包括脱氢加水再脱氢。脱氢产生FADH2【不饱和脂肪酸不脱氢,通过烯酰-CoA异构酶。多不饱和脂肪酸通过2,4-二烯酰CoA还原酶,多消耗一个NADPH【而不是NADH】。】,不通过复合体II进入呼吸链【通过电子传递黄素蛋白、铁硫蛋白和CoQ】。降糖氨酸是这一步的抑制剂。可以产生代谢水,对于骆驼等生物很重要。软脂酸可以产生106个ATP。
- 奇数脂肪酸产生丙酰CoA,转变为琥珀酰CoA【最后一步自由基催化,需要脱氧腺苷钴胺素。缺少时产生甲基丙二酸抑制糖异生,血糖降低。】。
- 过氧化物酶体发生的β氧化【超长链脂肪酸。通过D类ABC转运蛋白主动运输而不是CPT,不需要肉碱】。脱氢由氧化酶催化【而不是脱氢酶】,还原氧气产生过氧化氢,不产生ATP。对短链脂肪酸不作用,需要肉碱出去,再进入线粒体。
- α氧化如植烷酸。需要活化,但不产生ATP。除了线粒体和过氧化物酶体,还可以发生在ER。缺陷导致Refusm病,症状包括视觉和神经。
- ω-氧化发生在末端甲基。不需要活化。
- 酮体产生在肝细胞的线粒体基质。丙酮主要通过肺呼出体外。肝细胞没有利用酮体的酶——酮酰CoA转移酶和乙酰乙酸硫激酶。
- 植物脂肪酸合成发生在质体。从头合成需要引物【乙酰CoA或丙酰CoA】。
- 线粒体内膜柠檬酸-丙酮酸穿梭系统,转移乙酰CoA,需要柠檬酸裂合酶。细胞能量状态高,异柠檬酸脱氢酶活性降低,柠檬酸离开细胞。
- 脂肪酸合酶是多功能酶同源二聚体。
- 脂肪酸合成产生软脂酸。其他的脂肪酸通过延伸和去饱和得到。延伸可以发生在ER和线粒体基质。去饱和在sER。哺乳动物的去饱和不能发生在编号高于9号的碳。亚油酸和α-亚麻酸必需。
- 丙二酸单酰CoA抑制CPTI。ACC调节包括别构引起单体和多聚体互变【多聚体有活性。柠檬酸激活,软脂酰CoA抑制。】和磷酸化【胰高血糖素PKA和AMP的AMPK】。量变调节通过PPAR【过氧化物酶体增值激活受体】。胰岛素通过USF和SREBP【固醇应答元件结合蛋白】刺激脂肪酸合酶表达。瘦蛋白和多不饱和脂肪酸抑制SREBP和脂肪酸合酶。
- 肝脏中ACC1合成丙二酸单酰CoA,合成脂肪酸。心脏和骨骼肌中ACC2合成丙二酸单酰CoA,用来抑制CPT-I,在剧烈运动时ACC2磷酸化被抑制。
胆固醇代谢
- 胆固醇合成主要在肝细胞。位于细胞质基质和sER【大多数步骤。HMG-CoA合成后】。
- 胆固醇合成的前两步和酮体合成的步骤一样,但发生在细胞质基质【酮体合成在线粒体基质】。
- HMG-CoA还原酶是限速酶,需要两分子NADPH把酰基还原。斯达汀抑制。
- 甲羟戊酸合成异戊二烯PPi【异构为二甲烯丙基PPi】,需要脱羧。此后合成牻牛儿PPi,法尼PPi。消耗NADPH生成鲨烯。鲨烯通过固醇载体蛋白转移到内质网。通过环氧鲨烯【鲨烯单加氧酶,重要的人工调控位点,不会影响其他萜类代谢】、羊毛固醇等变为7-脱氢胆固醇和胆固醇。
- 胆固醇需要和脱辅基脂蛋白结合。按密度从低到高,分别是CM,VLDL,IDL,LDL,HDL。单层磷脂。
- 脂蛋白可以
- 作为配体参与内吞,如ApoB-100被LDL受体识别。
- 调节脂代谢酶活性。如CM和VLDL表面的ApoC-II激活【脂蛋白脂肪酶】LPL。HDL表面的ApoA-I激活LCAT【卵磷脂:胆固醇脂酰基转移酶】。
- 诱发残体清除。ApoE诱导血液中VLDL和CM残体清除。
- 食物到肝细胞:CM。CM通过淋巴入血液。主要成分是脂肪。含有ApoB-48。后来得到ApoC-II被水解,变为残体。再得到ApoE,被肝细胞吸收。
- 小肠上皮细胞吸收胆固醇需要NPC1L1膜蛋白感受固醇,植物固醇被ABCA5/8运回消化道。通过内质网腔的MTTP【微粒体脂肪转移蛋白】进入内质网,缺陷导致β脂蛋白血症。
- 肝细胞到肝外细胞:VLDL,IDL,LDL。LDL含有ApoB-100。被LDLR识别。LDLR缺陷导致LDL水平增加,动脉粥状硬化【坏胆固醇。沉积、氧化、吞噬、裂解、再吞噬,炎症。抗氧化剂VC和VE可以防止LDL氧化。】。PCSK9【前蛋白转移酶枯草溶菌素9】降解LDLR,是人工调节热点。
- 肝外细胞到肝:HDL。胆固醇从外周细胞到HDL需要ABC1、ApoA-I和LCAT。CETP催化胆固醇酯在HDL和VLDL的转移。好胆固醇。SR-B1【清道夫受体】帮助HDL上胆固醇的清除。人工制作插入ApoA-I的脂质体可以用于治疗。
- 胆固醇代谢需要酯化,合成胆汁酸。酯化可以由ACAT催化,在ER,供体脂酰CoA。LCAT催化在血清,和ApoA-I结合。供体卵磷脂。
- 肝细胞合成胆汁酸。需要ABCC2,缺陷导致Dubin-Johnson综合征和黄疸。肠肝循环。排除到体外的胆汁酸可以看作胆固醇清除的唯一通道。初级胆汁酸包括胆酸和鹅脱氧胆酸。次级胆汁酸包括7-脱氧胆酸和石胆酸,是消化道内细菌转化来的。
- HMG-CoA还原酶通过磷酸化、降解和基因表达调控被调节。去磷酸化形式具有活性,由AMP下游的AMPK催化磷酸化。PKA下游的HMG-CoA还原酶磷酸酶去磷酸化。降解通过固醇感应结构域。
氨基酸代谢
-
LAAO和DAAO不是脱氨基的主要蛋白,为黄素蛋白,存在于动物和细菌的过氧化物酶体。
-
谷氨酸脱氢酶位于线粒体基质。是一种同时可以用 NAD+ 和 NADP+ 作为辅酶的酶。通过ADP-核糖基化抑制【可以用于调节胰岛素分泌】。
-
Ser和Thr 可以通过脱水酶脱氨。
-
Thr,Pro,Lys 不能进行转氨基反应。
-
联合脱氨基是主要的氨基代谢方式。
-
合成一个Asn需要两个ATP【PPi】,合成一个Gln需要一个ATP。
-
Gln作为代谢枢纽。GS【谷氨酰胺合成酶】的调控:原核生物九种别构抑制剂,腺苷酸化抑制。哺乳动物只有别构调节,只有酮戊二酸激活。
-
CPS-1 CPS-2 肝细胞线粒体基质 大多数细胞的细胞质基质 尿素循环 嘧啶核苷酸合成 NH3提供氨基 Gln提供氨基 -
尿素循环的调节:NAG【N-乙酰谷氨酸】的别构激活。——细菌鸟氨酸合成的一种中间物。
-
尿素循环一共消耗4分子 ATP。
-
尿素循环发生在肝细胞的胞质 和 线粒体。
核苷酸代谢
-
嘌呤环从头合成的原子来源:1天,28佳人,6探无应,服用39胃泰加骨鲜汤,745肝。
-
PRPP还能参与某些核苷酸类辅酶【NAD,NADP】和某些氨基酸【His,Trp】的合成。
-
PRPP合成酶的抑制剂:ADP,2,3-BPG,AMP,GMP,IMP
-
Gln:PRPP酰胺转移酶 将核糖 从 α构型转变为β构型。是限速步骤。调控通过 有活性的单体 和 无活性的二聚体之间的转变。IMP,GMP,AMP抑制。PRPP促进。
-
嘌呤核苷酸从头合成过程中的多功能酶:【Gln:PRPP酰胺转移酶,2甲酰基转移酶,4合成闭环的合成酶】,【5羧化酶(分子吸尘器,不需要ATP就可以羧化,因为下一步很容易发生),6转移Asp的合成酶】,【8甲酰基转移酶,9次黄苷酸合酶】
-
IMP生成AMP需要GTP。IMP生成GMP需要ATP。
-
骨骼肌细胞中存在嘌呤核苷酸循环。利用AMP和IMP的转化,把Asp变为延胡索酸,加速TCA循环。心肌细胞缺氧可以通过HIF-1α将延胡索酸作为呼吸链末端电子受体。并且肌肉细胞谷氨酸脱氨酶活性低,主要通过腺苷脱氨酶和嘌呤核苷酸循环给氨基酸脱氨。
-
嘧啶核苷酸先合成嘧啶环,最后再形成糖苷键。有的反应发生在线粒体【乳清酸脱氢酶位于线粒体内膜】。
-
嘧啶核苷酸从头合成的原子来源:3姑2叹4天。
-
CPS-2是真核生物嘧啶核苷酸合成的限速酶。细菌则是ATC【氨甲酰天冬氨酸的形成】。CPS-2被UDP,UTP抑制,被PRPP激活。EGF磷酸化使得CPS-2被抑制但同时对PRPP更加敏感。
-
嘧啶合成中的多功能酶:前三步,后两步。
-
胞苷酸需要UMP先转化为UTP,再通过CTP合成酶合成CTP。
-
嘌呤核糖转移酶包括 APRT 和 HGPRT,需要焦磷酸。嘧啶核糖转移酶则主要通过 核苷磷酸化酶 和 核苷激酶。
-
核苷单磷酸激酶对戊糖没有特异性。核苷二磷酸激酶对碱基和戊糖均没有特异性。
-
核苷酸还原酶涉及自由基的产生。第一种分布最广,底物是NDP【NDP还原酶】,涉及酪氨酸自由基,需要双铁氧桥。第二种需要脱氧腺苷钴胺素的自由基【β氧化奇数脂肪酸中,甲基丙二酸单酰CoA变位酶也需要】,在蓝细菌中。
-
羟基脲和麦芽酚镓可以抑制NDP还原酶。羟基脲通过去除自由基,麦芽酚镓通过取代活性中心的铁原子。
-
NDP还原酶的调控机制:
-
嘌呤核苷酸的 1-磷酸核糖 通过 磷酸脱氧核糖醛缩酶 被裂解成为 3-磷酸甘油醛和乙醛。
-
尿酸的形成——黄嘌呤氧化酶需要金属Mo【含钼酶包括黄嘌呤氧化酶、固氮酶、硝酸还原酶、亚硫酸氧化酶、醛氧化酶、黄嘌呤脱氢酶等】。
-
嘌呤碱基的分解:尿酸——灵长类、鸟类、爬行类、昆虫。尿囊素——大多数哺乳动物。尿囊酸——硬骨鱼。尿素——两栖动物、软骨鱼。铵根——海洋无脊椎动物。
-
嘧啶碱基的分解:氧化直到二氢尿嘧啶开环,再脱羧脱氨基。最后形成丙二酸单酰CoA或甲基丙二酸单酰CoA。氨基异丁酸也可以通过尿液排出【因为只有DNA含有T,T降解产生氨基异丁酸,可以用氨基异丁酸代表DNA代谢水平】。
-
痛风是尿酸堆积过量——PRPP合成酶、Gln:PRPP合成酶的上调,HGPRT的下调。别嘌呤醇治疗,是黄嘌呤氧化酶的自杀抑制剂。严重联合免疫缺陷病——腺苷脱氨酶。白细胞受到影响最大。Lesch-Nyhan综合征——HGPRT。乳清酸尿症——多功能酶【乳清酸磷酸核糖转移酶和OMP脱羧酶】。使用尿嘧啶或尿苷治疗,生成UMP抑制CPS-2.
-
叶酸类似物影响嘌呤核苷酸的合成,如氨基蝶呤,磺胺药。
-
谷氨酰胺类似物包括重氮丝氨酸和重氮氧正亮氨酸。
-
6-巯基嘌呤用于儿童白血病的治疗。5-FU产生5-FUMP,是胸苷酸合酶的自杀抑制剂。
-
AZT,ddI用于阻断HIV反转录。araC用于治疗急性白血病。
-
乳清苷酸脱羧酶也是一个调控位点,被UMP抑制。
分子生物学
DNA复制
-
识别 复制起始区 的蛋白质:
细菌 古菌 真核生物 DnaA Orc1/Orc6 (Orc1-Orc6) -
几乎所有的 DNA聚合酶都涉及 2 个 Mg++【一个是本来就有的,一个是 dNTP带进来的】 和 3 个 Asp。
-
大肠杆菌的 DNAP I的3'-外切酶用来校对,5'-外切酶用来切除引物。
-
大肠杆菌 DNAP III 的结构很复杂,是大肠杆菌复制的主要酶。核心酶—— α【聚合酶】,ε【3'-外切酶】,θ。pol III’——核心酶 + τ【装配】。β——滑动钳【真核生物中的 RFC和 PCNA 是它的类似物】。钳载复合物——gama,delta,delta’,psi…
-
真核生物有 DNAP A【引发酶】,DNAP B,DNAP C,DNAP D【后随链】、DNAP E【前导链、修复】
-
DNAP A 没有 3’-外切酶活性。RPA 和它结合稳定它。
-
真核生物中的 PCNA 是 大肠杆菌中滑动钳的类似物。3个PCNA组成滑动钳 。RFC帮助它组成滑动钳。
-
大多数解链酶优先和 单链 结合。
-
DNA拓扑异构酶通过Tyr 两次转酯反应 催化。topo I 切断单链【其中topo I可以分为topo IA和topo IB.历史上,IA型拓扑异构酶被称为原核拓扑异构酶I,而IB型拓扑异构酶被称为真核拓扑异构酶。从功能上来说,这些子类执行非常专门的功能。topo IA只能松弛负超螺旋DNA,而 topo IB可以引入正超螺旋,在DNA复制后分离子代染色体的DNA,松弛DNA。】,而 topo II 切断双链。参与DNA复制的主要topo II。细菌的旋转酶属于 topo II。环丙沙星和新生霉素抑制旋转酶。
-
引发酶:
| 细菌 | 真核生物 |
|---|---|
| DnaG | DNAP A |
| 11nt | 7-10nt |
- 切除引物的酶:
| 细菌 | 真核生物 |
|---|---|
| DNAP I,RNAse H | FEN 1,RNase H1 |
-
DNA连接酶——第一类利用 ATP,第二类使用 NAD+【细菌】。都涉及 Lys 的 酶-AMP中间物。最后释放出 AMP和PPi或 NMN。
-
维纳综合征:端粒酶活性低。
-
DNA的复制以 复制子 为单位。
-
细菌,古菌,真核生物 DNA 复制的主要蛋白质和酶的比较:
| 蛋白质和酶 | 细菌 | 古菌 | 真核生物 |
|---|---|---|---|
| 起始 | DnaA | Orc1/Orc6 | ORC |
| 解链 | DnaB | MCM | MCM复合物 |
| 解链酶转载物 | DnaC | Orc1/Orc6 | Cdt1/Cdc6 |
| DNAP | C | B | B |
| 滑动钳 | DNAP III β | PCNA | PCNA |
| 滑动钳转载物 | γ | RFC | RFC |
| DNA连接酶 | NAD+ | ATP | ATP |
| 切除引物 | DNAP I/RNase H | RNaseH/FEN1 | RNaseH/FEN1 |
| 端粒酶 | 无 | 无 | 有 |
- 细菌DNA复制开始于 OriC的识别。OriC包括:
-
9bp直接或反向重复,A盒【DnaA识别】
-
13bp直接重复,DUE,DNA解链元件【最先发生解链】
-
GATC甲基化位点【Dam甲基化调控复制起始】
-
CTG序列【被引发酶 DnaG 识别】
-
细菌DNA复制的起始需要 DnaA【复制起始蛋白。在HU和IHF的帮助下,识别OriC的 A盒【HU为非特异性,IHF为特异性】,解链DUE】,DnaB【解链酶】,DnaG【引发酶,在PriA,PriB,PriC帮助下,结合 CTG序列。11nt。】
-
后随链 之所以能够向前行进是因为形成了 突环 结构。
-
细菌DNA复制的终止,具有 Ter位点,被Tus蛋白结合。Tus蛋白是 解链酶DnaB 的抑制剂。具有作用 极性,如果DnaB碰到Tus蛋白β股的一面,就不能通过。子代DNA之间的 分离 需要通过 拓扑异构酶IV 和 XerCD。
-
滚环复制发生在 M13噬菌体和一些小的质粒,但在真核生物也有。A蛋白 同时 具有内切酶和复制起始蛋白的活性。不需要RNA引物,直接用游离的 DNA 3’-OH 作为引物。
-
D环复制发生在线粒体和叶绿体,腺病毒。分为重链和轻链。解链由TWINKLE催化。RNA引物由POLRMT【哺乳动物】或者TWINKLE【某些非后生动物】合成。引物是接近基因组全长的 mRNA,需要 RNase MRP 剪切后成熟。后随链也是不连续合成。前导链和后随链合成时间上不同步。
-
真核生物具有多个复制子。它们不一定同时启动。
-
真核生物具有 Orc【复制起始蛋白。】、Mcm【解链酶】、DNAP A【引发酶,RPA作为SSB帮助结合。7-10nt。】,DNAP D、DNAP E、PCNA【滑动钳,RFC帮助形成】、拓扑异构酶I【释放超螺旋张力】、拓扑异构酶II【分开连环体】、FEN1、RNaseH1【内切核酸酶。不能切断单个核糖核苷酸】。
-
真核生物DNA复制的开始。需要起始区识别蛋白复合物ORC的形成。pre-RC形成需要执照因子 Cdt1 和Cdc6 的帮助。此后Mcm加入,招募 DDK,CDK,Cdc45,GINS。DDK,CDK磷酸化活化Mcm。Cdc45-Mcm-GINS【CMG复合物】进一步增强 Mcm 活性。DNAP A引发酶加入,形成有活性的复制叉复合物。
-
真核生物DNA复制的延伸。需要 Ctf4/1,RPA【作为SSB】。DNAP A合成7-10nt的引物,随后合成20-30nt的DNA。解链——Mcm。引物水解——FEN1,RNaseH1。拓扑异构酶I——清除正超螺旋,和拓扑异构酶II作用相反。
-
真核生物DNA复制的终止。裂殖酵母,复制终止位点【RTS】,其中复制叉障碍物【RFB】具有极性。
-
拓扑异构酶II,防止终止区DNA断裂和重组。Ia,帮助姐妹染色单体分离。
-
古菌同时具有 DNA旋转酶 和 组蛋白。组蛋白形成四聚体核心。极端高温物种具有负旋转酶,引入正超螺旋。
-
细菌 DNA 复制起始的调控。GATC 位点被 Dam 甲基化,使得抑制蛋白 SeqA 和 OriC 解离。复制完的细菌DNA半甲基化。此外这个事件也参与了DNA的 错配修复。MutS 结合DNA并募集 MutL,激活内切酶 MutH。
DNA转录
-
RNA聚合酶没有3’-外切酶活性,却具有潜在的内切酶活性【可以在延伸阶段,在 GreA,GreB 的帮助下切除3’-端核苷酸,使得3‘-OH回到活性中心,解除暂停状态】。所以并不是没有校对机制。
-
RNAP 的催化机制也涉及 2个Mg++ 和 3个Asp。
-
转录的起始,RNAP 会多次催化无效转录,DNA会在它的活性中心产生褶皱,以便在无效转录时仍然可以和RNAP结合。
-
RNAP 的反应速率很低。50nt/s。
-
原核生物的 RNAP:
α β β’ ω σ 二聚体,核心酶的组装 转录 和DNA非特异结合 β‘的分子伴侣 启动子的识别 -
原核生物的 σ因子是可变的,起到了真核生物中转录因子TF的作用。
-
细菌RNAP的抑制剂是 利福霉素 和 利链霉素。【旋转酶的抑制剂是环丙沙星 和 新生霉素。】
-
RNAP I RNAPII RNAP III rRNA(除了5SrRNA) mRNA,有帽snRNA和snoRNA.miRNA,lncRNA 小RNA.包括tRNA,5SrRNA,无帽snRNA,snoRNA.7SLRNA,7 SK RNA,RMP RNA,端粒酶 RNA,少数lncRNA -
植物细胞还具有 RNAP IV 和 RNAP V,转录长的 lncRNA,参与RNA引导的DNA甲基化。
-
真核生物RNAP II 的最大亚基和细菌 β‘ 同源。具有CTD【羧基端结构域】。CTD 磷酸化 使得转录进入 延伸,对后加工很重要。
-
α-鹅膏蕈碱 抑制真核生物转录【RNAP II最敏感,RNAP I其次,RNAP III 最不敏感】,放线菌素D 能够插入GC碱基对【但是对RNAP I最敏感。因为rDNA具有较多的GC碱基对】。
-
古菌需要转录因子,但没有CTD。
-
确定启动子的方法有电泳涌动变化分析,DNA酶I足迹法分析。
-
细菌启动子包括 -35区【被σ70识别。TTGACA】 和 -10区【Pribnow Box.TATAAT】。rDNA还具有增效元件【-40 to -60】
-
启动子的一致序列是统计结果。
-
细菌RNAP形成转录起始复合物是限速步骤。细菌RNAP 先非特异性结合,然后扫描到 -35区,形成封闭复合物【主要以静电引力结合】。变构形成开放复合物【也就是转录起始泡,还可以以氢键结合。限速步】。
-
细菌RNAP起始阶段涉及无效转录。DNA分子形成褶皱保持对细菌RNAP 的结合。
-
启动子清空 由σ因子解离决定。
-
NusG和RfaH在转录中形成滑动钳结构。转录过程中,转录泡为 17bp,有 8 bp片段形成 A型双螺旋的杂交双链。若在延伸时暂停【NTP短缺】,需要 GreA,GreB 解除暂停。【在GreA,GreB的帮助下,细菌RNAP 的 内切酶 活性激活,切除3’-端核苷酸,使得3‘-OH回到活性中心,解除暂停状态】
-
转录的终止。不涉及ρ因子的终止机制需要终止子——U序列【形成茎环后U序列氢键强度小,解离】,和U序列上游的富含GC碱基对的 茎环。涉及ρ因子的终止机制需要ρ因子【具有解旋酶和ATP酶活性,和FoF1ATP合酶同源,结合变构】,rut【ρ因子结合位点】,3’-端也具有茎环结构,但 不 具有U序列。
-
真核生物RNAP I。
-
RNAP I的启动子。包括 CP【核心启动子】和 UCE【上游控制元件】,它们也同源。
-
RNAP I的转录因子。哺乳动物基础转录因子包括UBF【UCE结合因子】,SL1或TIF-IB【选择因子1或转录起始因子-IB。包括TBP【TATA盒结合蛋白】和3个TAF【TBP相关因子】。定位因子引导 RNAP I】。此外还可能具有 TIF-IA和TIF-IC。
-
RNAP I 转录的终止:哺乳动物终止子为Sal盒。可以招募Rnt1和Ran1【RNase】。鱼雷模型——Rnt1内切剪开正在转录的rRNA。Ran1外切追逐RNAP I。在Sen1【解链酶】的作用下,RNAP I 被释放。
-
真核生物RNAP III。
-
RNAP III的启动子包括:
-
外部启动子:TATA盒,PSE【近序列元件】,DSE【远端序列元件】。涉及7SK RNA,7SL RNA,U6 snRNA.
-
内部启动子:启动子位于基因内部。涉及tRNA【A盒,C盒,中间元件】,5SrRNA【A盒-D环,B盒-TφC环】,腺病毒的 V A RNA。
-
RNAP III的转录因子包括TF IIIA【只有5SrRNA的转录需要】,B【定位因子。包括TBP,BRF1【TF IIIB相关因子】,TF IIIB‘’】,C【组装因子,和A盒、B盒结合】。
-
tRNA转录的起始和延伸。TFIIIC结合A盒、B盒。5SrRNA转录的过程。TFIIIA结合启动子。招募TFIIIC。
-
基因外启动子的转录。需要Octa,STAF【转录因子】和DSE。需要SNAPC【snRNA激活蛋白复合物】结合PSE。它们也是RNAP II催化snRNA需要的。
-
真核生物RNAP II。
-
RNAP II的核心启动子包括 TATA盒,Inr【起始子】,BRE【TFIIB 识别元件,是转录因子TFIIB的识别序列】,MTE【模体10元件,依赖Inr】和DPE【下游启动子元件,依赖Inr】。TATAbox 和 Inr属于招募和定位元件,决定转录起点。RNAP II的调控元件包括UPE【上游邻近元件】和UIE【上游诱导元件】。
-
RNAP II的转录因子包括 TFIID【包括TBP和TAF。识别和结合核心启动子。招募其他转录因子。调节蛋白目标。具有激酶活性、组蛋白乙酰转移酶和泛素激活酶/结合酶活性。】,TFIIA【结合TBP。去除TBP抑制。稳定TBP和TATA盒的结合。】,TFIIB【招募RNAP II 和 TFIIF】,TFIIF【启动子的解链。无TFIIB时去磷酸化CTD】,TFIIE【招募TFIIH】,TFIIH【模板解链。形成第一个磷酸二酯键。磷酸化CTD,启动子清空】。转录延伸因子【包括NELF,DSIF,P-TEFb,TFIIS。NELF和DSIF暂停转录【和加帽反应有关】。P-TEFb解除转录暂停。TFIIS类似细菌GreB蛋白解除转录暂停】。RNAP II 转录暂停的解除需要TFIIS【C端结构域刺激RNAP II的内切酶活性】。
-
RNAP I RNAP II RNAP III CP【核心启动子】+UCE【上游控制元件】 核心启动子:TATA盒+Inr+BRE【TFIIB】+MTE+DPE;UPE;UIE 外部启动子:TATA盒+PSE+DSE【7SKRNA+7SLRNA+U6snRNA】;内部启动子【tRNA+5SrRNA+VARNA】 UBF+SL1+TIF-IB【TBP+TAF】 TFIID【TBP+TAF】+TFIIA+TFIIB+TFIIF+TFIIE+TFIIH;转录延伸因子NELF,DSIF,P-TEFb,TFIIS。 外部:Octa+CTAF+DSE+SNAPC;内部:TF IIIA,B【TBP+BRF1+TF IIIB'】,C NELF+DSIF+P-TEFb+TFIIS -
RNAP II的介导蛋白包括 SRB蛋白【和CTD直接结合】,SWI/SWF蛋白【破坏核小体结构,促进染色质的重塑】。
转录后加工
- 细菌的mRNA很少经过后加工。一个极少发生的例外是,某些细菌mRNA具有内含子。但不少细菌mRNA具有多聚A尾。和真核生物相比,细菌的多聚A尾巴较 短,为15-50nt,且多为 降解的 信号。真核生物中多聚A尾巴较长200nt,并且功能也不一样。
- 所有生物均有rRNA以多顺反子的形式被转录。细菌有的tRNA夹在rRNA中。
- 细菌rRNA的加工包括剪切、修剪【RNaseIII,D,F,P,M16,M5】和核苷酸的修饰。剪切为粗加工,内切;修剪为细加工,外切。其中16SrRNA和23SrRNA需要通过 茎环 结构被识别。修饰包括核糖的2‘-OH甲基化和假尿苷化。
- 细菌的tRNA加工包括剪切、修剪、核苷酸的修饰、CCA的加入和剪接。剪切和修剪产生的有前导序列【RNaseP,5'-剪切】和拖尾序列【若 无CCA,RNase Z在3’-剪切,此后加上CCA。若 有CCA,RNaseF,D】【uzi又发电】。核苷酸的修饰包括TφC。CCA的添加需要CCA添加酶或tRNA核苷酸转移酶,不需要模板。少数细菌的tRNA需要剪接,为第一类内含子。
- 真核生物的mRNA加工包括5‘戴帽,3’加尾,甲基化、剪接和编辑。
- 聚合酶II最大亚基上的CTD的 Ser2磷酸化 加尾,Ser5磷酸化 加帽。
- 5’戴帽——mRNA的帽子结构是5‘,5’-三磷酸酯键的甲基化鸟苷酸【m7Gppp】。可以按照其后的核糖甲基化与否分为0,1,2型。加帽反应是共转录的【延伸阶段,TFIIH磷酸化CTD的Ser5,通过DSIF和NELF暂停转录。暂停时加帽的酶识别Ser5加帽。P-TEFb磷酸化Ser2,解除暂停】。任何由pol II转录的RNA都具有帽子,除非后来被切除了。
- snRNA也具有帽子。是2,2,7三甲基化鸟苷酸帽子,后两次甲基化发生在细胞质。
- 3’端加尾——250nt。和细菌、古菌、线粒体和叶绿体相比长,稳定,且功能不一样。最重要的加尾信号是AAUAAA。加尾前需要剪切。参与加尾的蛋白质包括CPSF【特异性因子】,CstF【剪切刺激因子】,CF【剪切因子】,PAP【poly A聚合酶】,PABP【类似于SSB,结合短的poly A帮助其延伸】。其中PABP有PABPN1和PABPC1,分别位于细胞核和细胞质,结合在转录中和转录后。
- 加尾在哺乳动物线粒体中可能引入一个终止密码子。很显然引入的这个终止密码子可能是UAA或者UGA。
- 甲基化——m6A,GAC序列中。和细菌DNA复制对比:细菌Dam催化GATC序列中A的甲基化,刺激复制发生。
- 剪接——剪接就是切除内含子,拼接外显子。本质是两次转酯反应。剪接信号包括5‘-剪接点、3’-剪接点和分支点。剪接点满足 GU-AG规则。分支点的一个A很重要。需要的蛋白质有U1,U2,U4,U5,U6。U1和5’剪接点结合,U2和分支点,U5连接两个外显子。U6可以取代U1和5‘剪接点处结合。
- 第一次转酯发生在5‘-剪接点和分支点,需要U6【剪接点】和U2【分支点】结合拉动,通过U4/U6被U2/U6取代。第二次转酯反应发生在5‘-剪接点和3’剪接点,需要U5拉动两个外显子。
- 剪接体的拼装依次经过 A复合物,B复合物,C复合物,P复合物。U1,U2结合产生A复合物,U2取代了原本结合分支点的蛋白BBP【分支点结合蛋白】。U4/U6,U5进入,U1离开,形成B复合物,U4/U6被U2/U6取代。导致【U6】剪接点和【U2】分支点靠近,第一次 转酯反应发生,C复合物形成。U5把两个剪接点拉近,第二次 转酯反应发生,P复合物形成。
- U1 是 真核生物转录组的保护神。可以防止转录【而不仅仅是剪接】提前终止。通过保证加尾反应只在转录完成后发生。
- 次要剪接途径服从 AU-AC规则。U11,U12,U4atac,U6atac分别取代了U1,U2,U4,U6。值得注意的是U5是共用的。
- 反式剪接 发生在 叶绿体 和锥体虫、线虫等生物的核基因组中。秀丽隐杆线虫很多基因具有操纵子结构,并为多顺反子,需要通过反式剪接产生单顺反子,同时还要加上具有帽子结构的前导序列才能保证翻译。
- 复杂的多细胞生物在很大程度上依赖选择性剪接。而酵母则不是。
- 编辑——编辑包括转换、颠换和核苷酸数目的增减。转换和颠换需要通过脱氨酶。ADAR和CDAR【AR代表的是act on RNA,作用在RNA】分别催化了C-U和A-I(G)的转变。C-U的例子是载脂蛋白ApoB48和ApoB100。数字小的那个更短,通过引入了一个终止密码子。A-I在真核生物中甚至更为广泛,它的缺陷会导致遗传性对称性色素异常症,它的例子包括谷氨酸受体和它本身。ADAR2是一个自催化的酶。
- 核苷酸数目的增减则需要gRNA的参与。它的作用方向是3’-5‘。它的作用对象是U。它可以切除U序列,也可以增添U序列,通过尿苷酸转移酶或者尿苷酸外切酶。
- 真核生物的rRNA修饰需要snoRNP的帮助。注意这不是snRNP而是snoRNP。它的功能比较丰富,可以 剪切,但更多地是参与 修饰。C/Dbox 结构原件可以引入2‘-OH的甲基化,而 H/ACAbox 可以引入假尿苷。RNaseMRP参与了5.8SrRNA的剪切,同时它还在线粒体mtDNA复制中,通过把一个和基因组差不多大的巨大RNA剪切产生引物。
- 真核生物的tRNA修饰和细菌大致相同。一个显著的区别是可能具有小的内含子。这些内含子的去除会产生特别的2’-3’环磷酸【不是3’-5’环磷酸】,需要水解产生3’-OH才行。不仅如此,磷酸基团总是要连接到5’-OH的。它需要一个GTP来把磷酸基团转移到5‘-OH上。【而在细菌中,这一切都是自催化的。它是一个第一类内含子。】另一个显著的区别是真核生物的tRNA绝对没有CCA。和细菌一样,需要被RNaseZ切除之后加上。
- 古菌基本上和细菌更为相似。一个独特的地方是 tRNA可以进行 编辑,通过 CDAT【AT表示的是act on tRNA,作用在tRNA上】。另一个独特的地方是rRNA的剪接机制和真核生物完全不同,倒是和tRNA的剪接机制相同
mRNA的翻译
-
哺乳动物成熟的红细胞不能翻译蛋白质。
-
细菌核糖体小亚基【16SrRNA】识别mRNA 5’端的起始密码子。大亚基【23SrRNA】具有转肽酶活性。真核生物 28SrRNA 和 5.8SrRNA 和细菌 23SrRNA 有同源关系,可能为同一个基因断裂得到。
-
古菌的核糖体在大小和组成上类似于细菌。在一级序列上类似于真核生物。
-
ORF【开放阅读框】是指起始密码子到终止密码子的区域。ORF不一定是CDS。
-
tRNA的个性包括正效元件和负效元件。经常出现在受体茎和反密码子环。
-
起始tRNA和tmRNA是特殊的tRNA。
-
aaRS【氨酰tRNA合成酶】消耗两分子ATP【PPi供能】。催化氨酰AMP合成。包括两类。
-
第一类:单体酶,Rossman折叠。个性包括受体茎 和 反密码子环。结合在受体茎 小沟。先 连接到2’-OH,后转酯到3’-OH.
-
第二类:寡聚酶,签名模体。个性 不 包括反密码子环。结合在受体茎 大沟。直接 连接到3‘-OH。
-
第一类 第二类 单体酶 寡聚酶 Rossman折叠 签名模体 受体茎和反密码子环 只有受体茎 结合在小沟 结合在大沟 先连接到2’-OH 直接连接到3’-OH
-
-
辅助蛋白因子包括 IF【起始因子】,EF【延伸因子】,RF【释放因子】。细菌还具有RRF【核糖体循环因子】。
-
起始密码子有AUG,UUG,GUG。终止密码子有UGA【可作Sec 】,UAG【可作Pyl】,UAA。
-
大多数第二位嘌呤的密码子编码亲水氨基酸。嘧啶编码疏水氨基酸。第三位碱基最不保守。
-
终止密码子后的第一个核苷酸。细菌多U,真核生物多G。
-
摆动法则和超摆动法则【线粒体】:【反密码子第一个碱基——密码子第三个碱基】
翻译后加工
重组DNA技术
- 细菌质粒的特点:共价闭环双链DNA;含有DNA复制起始区【严紧控制和松弛控制】;对宿主细胞的生存并不必须,通常有利于宿主在特定情况下生存。
- α互补、插入失活和蓝白筛选法——大肠杆菌质粒pUC18/19具有lacZ’【只能编码N端的146AA】,和宿主菌表达的C端肽链结合获得活性。在放入 IPTG 和 显色底物 X-Gal 时水解产生蓝色。序列插入后lacZ’的ORF失活,菌落白色。
- 噬菌体载体用来克隆 稍大的DNA片段【10-20kb】。
- BAC拷贝数受到 严紧控制。
- 动植物病毒作为载体需要引入质粒的复制起始区——穿梭载体。
- 导入载体的工具:RE,DNA连接酶,DNA聚合酶,逆转录酶,核糖核酸酶H【水解和DNA配对的RNA】,多聚核苷酸激酶【同位素标记】,S1核酸酶【水解单链】
- 引入细胞的方法:转化【冰冷CaCl2+热激;小DNA;转化率:超螺旋环状>带缺口开环结构>线性结构】;转染【两个意思。在细菌中为噬菌体侵染。在哺乳类中类似转化,DNA-磷酸钙共沉淀】;电穿孔;脂质体介导;弹道基因转移。
- 间接筛选法:核酸杂交、PCR、免疫化学、受体和配体、Southwestern/Northwestern印迹法【核酸结合蛋白质】、RE图谱分析法【酶切位点】、DNA序列分析法【最后鉴定】
- 有RNA质粒。
- 目的基因和载体的连接:
- 粘性末端:分离纯化后混合,低温退火即可连结。
- 平末端:削平【核酸酶S1】或补平【klenow酶】或RE再切,最后用T4DNA连接酶。
- 平末端和粘性末端:特别少见。只能通过一种方向插入载体。
- 利用cDNA文库,可以:确定基因的转录产物和翻译产物;表达不同的蛋白质;获得没有内含子的基因;体外转录mRNA;合成探针;基因突变分析;确定基因组序列中的基因;获得建立物理图谱需要的标签
- 转基因动物新霉素筛选。转基因植物卡拉霉素筛选,二元质粒系统——质粒1含有外源基因,质粒2含有vir将外源基因切出来整合到染色体上。
- 基因功能研究:基因敲除使用同源重组、转座子插入、基因编辑。基因敲减使用DNA修饰、人工设计核酶定向切割、RNAi、核酸酶H。显性负效突变。
- 寻找未知基因【核酸水平】:同源序列、基因标签、消减杂交【检测来自不同cDNA文库的异源双链,比对差异】,差异显示PCR【DD-PCR】,RNA随机引物PCR【RAP-PCR】,外显子捕获,和CpG岛有关的基数,噬菌体展示,酵母双杂交。
- PCR和DNA复制的区别:解链通过热变形;引物是人工合成;DNA聚合酶耐热。
- PCR过程:变性【95℃】、退火【50-65】、延伸【72】。原料:模板,耐热的DNA聚合酶,一对引物,4种dNTP,Mg++,缓冲液。
- 逆转录PCR【RT-PCR】和实时定量PCR【q-PCR】。
- RT-PCR,qPCR和Northern杂交:Northern杂交全面复杂不精确不灵敏,RTPCR灵敏不精确,qPCR精确。
- qPCR原理——淬灭集团和荧光基团,Taqman双标记探针技术。CT——阈值的循环数。
- 蛋白质随机突变——易错PCR【Mn++代替Mg++,降低一种dNTP浓度,使用缺乏校对活性的聚合酶、提高Mg++】】。基因混排——将不同物种的同源基因混合,消化,PCR,模板互换重组发生。
- 定点突变原理。
- 核糖体展示技术——体外转录、翻译,亲和层析蛋白质带着核糖体和mRNA,洗脱后对mRNA进行PCR。
- 研究蛋白质相互作用:免疫共沉淀、亲和层析、共价交联、FRET、酵母双杂交【通过将一个蛋白和DNA结合结构域连结,另一个蛋白和转录激活结构域连结。具体过程:选择载体,将蛋白基因插入载体,转染,筛选,活性检测】
- 研究蛋白质-核酸相互作用:电泳泳动变化分析、DNA亲和层析、DNA酶I-足印分析和染色体免疫沉淀【ChIP。甲醛固定,随即切割,通过抗体沉淀复合物,去交联即可PCR后测序DNA】
- 基因组编辑【机体内。重点是找到高度定向切割的核酸内切酶——Crispr-Cas9】通过DSB的 非同源末端连接【NHEJ】和 同源重组【HR】。基因敲除——NHEJ,插入、缺失。特异突变引入——HR,提高重组效率。定点转基因——HR。有缺陷的基因纠正——HR,模板正常,同源重组过程中替换需要纠正的基因上的缺陷。
- 基因组编辑的核酸内切酶:MGN、ZFN、TALEN酶自己去识别特定的碱基序列。Cas9只管切割,识别交给gRNA。CRISPR/CAS9系统——Cas9,crRNA,tractRNA。如何解链——通过小的 间隔序列邻近模体【PAM】,被切割DNA的蛋白Cas结合【而不是结合Cas的gRNA】,进而解链。必须依靠目标序列周围的PAM。Crispr含有回文重复序列和间隔区。
基因表达调控
- 亮氨酸拉链(英语:leucine zipper,亦称为亮氨酸剪刀)是蛋白质中一种常见的三维结构模体,常见于许多转录因子的DNA结合结构域,因此涉及基因的表达调控。亮氨酸拉链在真核生物和原核生物的蛋白中都有发现,但以真核生物居多。亮氨酸拉链是一种作为二聚体化结构域的超二级结构,且使相互平行的α-螺旋之间产生粘附力。一个亮氨酸拉链包含了多个亮氨酸残基,通常每七个氨基酸残基就出现一次,这形成了一条两性的α-螺旋,疏水区只在其中一侧。这个疏水区提供了二聚化的区域,使得基序可以像“拉链”一样拉起来。此外,疏水亮氨酸区域对于其结合到DNA是不可或缺的。亮氨酸拉链常见于许多转录因子的DNA结合结构域,因此涉及基因的表达调控。例如,c-fos与c-jun(AP1转录因子),还有myc家族的成员包括myc、max与mdx1,它们是正常发育的重要调控物。如果它们在一些部位发生过量表达或突变,则会产生癌症。这些蛋白质以二聚体(同二聚体或异二聚体)的形式与DNA发生相互作用,又被称为碱性拉链蛋白(bZips)。
分子生物学方法
-
Dual-Luciferase Reporter Assay(双荧光素酶报告基因检测)是一种常用的生物检测方法,它通常以萤火虫荧光素酶 (Firefly luciferase)为报告基因,以海肾萤光素酶 (Renilla luciferase)为内参基因。所构成的报告系统具有灵敏度高、动态范围广、应用灵活等优势,广泛用于基因调控、非编码RNA靶向互作等研究领域。
在实验中,萤火虫荧光素酶和海肾荧光素酶都无需修饰,只要正常转录即具有活性。其中海肾荧光素酶作为转染的内参,来减少孔间细胞数量和转染效率对实验结果的影响。通过特定的荧光检测仪检测荧光量来判断酶的活性,进而反映基因间的关系。
细胞生物学
细胞骨架
- 驱动蛋白家族的成员。kinesin-13为M-驱动蛋白,马达结构域位于肽链中部,结合使得微管解聚。kinesin-5是反向四聚体,在极性相反的微管之间滑动,介导中心体向两极分离。kinesin-4染色体定位。kinesin-1细胞器和mRNA运输。kinesin-2黑色素体和鞭毛内运输。
- 在动物中,几乎所有类型的细胞上都存在非活动性初级纤毛,但血细胞是一个明显的例外。大多数细胞只拥有一个,与具有活动纤毛的细胞相反,嗅觉感觉神经元是个例外,气味受体位于此处,每个细胞拥有大约十个纤毛。某些细胞类型,例如视网膜感光细胞,具有高度特化的初级纤毛。在内耳毛细胞上发现的 动纤毛 是特殊的非运动纤毛。它们拥有运动纤毛的 9+2 轴丝,但缺乏提供运动的内部动力蛋白臂。在外部动力蛋白臂的允许下,它们确实会在检测到声音后被动移动。
- 运动纤毛的功能很大程度上取决于维持纤毛周围液体的最佳水平。上皮钠通道(ENaC) 在呼吸道、输卵管的整个纤毛长度上特异性表达,显然充当调节纤毛周围液体的传感器。在脉络丛的上皮细胞上发现了活动的、多个、9+0 纤毛。在早期胚胎发育过程中发现了没有中央单线对 (9+0) 的活动纤毛。它们以结纤毛的形式存在于原结的结细胞上。节点细胞是造成双侧动物左右不对称的原因。虽然缺乏中央装置,但存在动力蛋白臂,允许结纤毛以旋转方式移动。该运动产生了胚胎外液向左流过节点表面的电流,从而引发了发育中胚胎的左右不对称。
细胞核和染色质
- 核纤层除了可以支撑外层核膜,还和染色质有联系,特别是异染色质。
- 核膜在普通光镜下难以分辨。相差显微镜下可以看出核膜的界限。电子显微镜下可以仔细观察
- 研究核孔复合体的经典方法主要有:树脂包埋切片方法、负染色技术、冷冻蚀刻技术。
- 核孔复合体可以分为环、辐、栓。
- 亲核蛋白具有NLS。富含Lys和Arg,还常常含有Pro。NLS可以存在于蛋白的不同部位,不被清除。亲核蛋白入核识别、转移依靠importin,释放依靠Ran-GTP。Ran-GTP存在于细胞 核,Ran-GDP存在于细胞 质。此外还有核输出信号NES。
- laminA和laminC是同一个基因的不同拼接体。laminA/C表达具有特异性,laminB则没有。异染色质更容易和核纤层结合。laminA是DNA断裂修复必须的。laminA的突变会导致H-G早老综合征,Werner综合征。laminB的突变会导致P-H综合征。
- 人类基因组中编码DNA比例为1%-1.5%。小卫星DNA可以用于DNA指纹技术,微卫星DNA可以用于构建遗传图谱。
- H4是最保守的组蛋白,不存在变体。H3其次,但具有CENP-A,H3.3等变体。H1最不保守,鱼类和鸟类的成熟红细胞中,H1被H5取代,有的生物比如芽殖酵母缺少H1。
- 研究DNA和非组蛋白作用可以利用 凝胶延滞实验。核小体的发现利用了微球菌核酸酶。
- 染色质的各级结构。一级结构核小体——200bp。146bp缠绕组蛋白八聚体1.75圈【压缩7倍】,20bp缠绕H1,连接DNA大约60bp。二级结构螺线管——每圈6个核小体【压缩6倍】。三级结构超螺线管——【压缩40倍】。四级结构染色单体——【压缩5倍】。总共压缩了8400倍。放射环模型认为螺线管形成的微带是染色体高级结构的单位。
- 当染色质处于松散结构,H3暴露出的游离巯基可以和 有机汞 亲和。利用有机汞亲和层析和二硫苏糖醇洗脱可以把活性染色质分离。
- 组蛋白 H1 的 磷酸化 主要发生在有丝分裂期间。确定核小体的方向。
- 脊椎动物尤其是哺乳动物比无脊椎动物的 甲基化 程度要高得多。基因组印记 只在哺乳动物中出现。
- 着丝粒可以分为 动粒结构域【进一步分为内板、中间间隔、外板、纤维冠。和内板相联系的染色质是和微管相互作用的位点】、中央结构域【主体。alpha卫星DNA】、配对结构域【内部着丝粒蛋白,染色单体连接蛋白】。
- 端粒和染色体在核内的空间排布有关。
- 端粒最终会形成一个D环或者T环,保护染色质的末端。
- 核型分析的三大技术:低渗处理技术、秋水仙素的应用、植物凝集素【刺激血淋巴细胞转化、分裂】。
- Q带AT亮,GC暗。G带相同。R带相反【Reverse】。T带染色端粒【吉姆萨和吖啶橙】。N带也叫做Ag-As染色带,用来染色NOR。除了T带,R带也常用于染色端粒部分。
- 多线染色体来源于细胞核内有丝分裂,处于永久间期。灯刷染色体来源于减数分裂的双线期,转录活跃。
- 核仁的结构在超薄切片电镜下,可以分为 纤维中心【rDNA储存】,致密纤维组分【rRNA转录】,颗粒组分【核糖体亚基前体颗粒储存】。其中电子密度最大的是致密纤维组分。颗粒组分是核仁的主要结构。纤维中心存在嗜银蛋白。核仁被染色质包围,
核糖体
- 原核生物翻译过程中需要的GTPase包括IF2,EF-Tu【eEF-1A】,EF-G【eEF2】,RF。
细胞通信和信号转导
- SH2结构域识别 p-Tyr,如Src,Grb2,Shc,STAT。PH结构域识别 肌醇磷酸,如Akt,Sos。SH3结构域 主要通过识别脯氨酸和酪氨酸激酶相互作用,如PI3K,RasGAP,Cdc24,cdc25
- c-Src不是CSK。CSK在C端磷酸化并抑制c-Src。
- MAchR释放beta,gama亚基使得K+外流,细胞超极化。
- 霍乱毒素激活Gs,百日咳毒素抑制Gi。
- Fura-2和Fluo-3是可以和钙离子结合的荧光染料。
- 心房钠尿肽的受体是受体鸟氨酸环化酶。
- PKC和钙离子结合后贴膜。
- 细胞因子包括白介素、干扰素、集落刺激因子CSF,促红细胞生成素Epo,生长激素、催乳素。Epo激活STAT防止红细胞前体细胞的凋亡。
- PI3K-PKB信号通路的作用是细胞生存,还可以促进胰岛素刺激的葡萄糖摄取和储存【上调GLUT4】。
- TGFbeta的受体分为RI,RII,RIII。其中 RIII 是 蛋白聚糖,富集TGFbeta给RI,RII。RI,RII是受体丝苏氨酸激酶。
- Wnt和Hh信号通路TF静息状态下被泛素化降解。NFκB和Notch涉及抑制物或者受体本身被切割激活。
细胞周期和细胞分裂
- 酵母细胞周期的特点:细胞周期属于封闭式,核膜不解聚,纺锤体位于细胞核内。芽殖酵母的纺锤体组装和 S期 DNA 复制同时进行。
- 高等植物没有中心体。并且胞质分裂由成膜体介导形成细胞板。
- 黏连蛋白cohesin 包括smc1/3, scc1/3;凝缩蛋白codensin 包括smc2/4。
- 中心体在 G1 开始复制,在 S 期完成复制,在 G2 分离,然后在前期形成星体。
- 星体包括 gama-tubulin、中心体蛋白等。动物形成有星纺锤体,植物形成无星纺锤体。
- Mad 和 Bub 在中期之前位于染色体动粒的外板上,一旦其和 MT 接触就从动粒上消失。Mad 将会抑制 APC。从而调控了染色体整列。
植物形态解剖
植物细胞
- 筛管分子没有细胞核,有的真菌和藻类多核。
- 有色体的色素聚集在有色体众多的脂球内。
- 初生淀粉粒在叶绿体中合成,后来转移到其他部位变成次生淀粉粒。
- 脂肪锇酸染色成黑色,蛋白质碘液染色成暗黄色。
- 鞣质就是单宁,本质是酚醛类聚合物。
- 动植物细胞的三大区别是大液泡、质体和细胞壁。
- 纹孔膜的本质不是细胞膜,而是初生壁和中层。
- 多网生长理论。认为微纤丝在长形细胞中微纤丝的排列方向最初是水平的,后来被拉伸为直向的。
- 植物细胞和动物细胞有丝分裂的三大区别是没有中心体、形成了胞间连丝和形成了细胞板。
植物组织
- 原形成层在茎尖排列成为原形成层束,在根尖并不排列成束,而是成为一个圆柱体型,位于根尖的中心。
- 原套原体学说。认为原套只进行垂周分裂,通常为两层,L1和L2。原体进行各种方向的分裂。
- 细胞学分区学说。可以分为中央区、周围区、肋状区。中央区维持分生细胞数量。周围区产生叶。肋状区产生髓。
- 角质层的外面通常还有蜡层。比如李子和葡萄果皮上的白霜。
- 表皮细胞一般没有叶绿体,但是保卫细胞具有叶绿体。细胞壁的微纤丝辐射排列是保卫细胞控制气孔开闭的决定因素。
- 双子叶气孔的类型可以分为不规则型、不等型、横列型、平列型。
- 细胞壁的颜色和栓质化没有关系,而是和鞣质有关系。
- 周皮和皮孔。皮孔的发生要早于周皮的发生,涉及皮层细胞产生补充细胞,并最终产生皮孔木栓形成层。最终撑破,露出来的补充细胞最终成为死细胞。皮孔木栓形成层是最早产生的木栓形成层,后来和别的木栓形成层联系越来越多从间断分布变成连续分布。因此周皮产生要晚于皮孔产生。
- 某些草本植物茎中的厚角组织特别发达,茎上带有坚硬的棱。临时装片可以表现出高折光度。但永久装片因为失水和薄壁组织差不多。
- 纹孔缘的有无是具缘纹孔和单纹孔的根本区别。尽管纤维细胞的单纹孔很抽象,但是没有纹孔缘。
- 协同生长和侵入生长。有没有改变相邻细胞之间原有的接触面。
- 麻类植物的韧皮纤维可以用来做纺织原料。
- 环纹导管和螺纹导管都具有一定的伸长能力,所以在植物发育的早期出现,因为他们不影响植物器官的生长。
- 90
植物的营养器官
- 叶可以分为三部分。叶片、叶柄和托叶。单子叶植物一般没有托叶,并具有单叶。豌豆的托叶很大,执行叶片的机能。
- 复叶有三出复叶、羽状复叶、掌状复叶等等。单身复叶是退化的三出复叶,柑橘具有。
- 单子叶植物并没有像双子叶植物那样典型的栅栏组织分化。栅栏组织仍然有良好的细胞间隙,但因为叶的横切面一般切的比较厚,所以看起来细胞间隙不显著。光合作用是它主要的机能 。
- 海绵组织的细胞间隙显著,排列疏松。但因为海绵组织细胞彼此之间接触面大,所以单位体积海绵组织和细胞间隙的接触面比栅栏组织和细胞间隙的小得多。气体交换是海绵组织的主要机能。
- 包围中脉的薄壁组织上下总是分布有两堆机械组织。在双子叶植物多为厚角组织,在单子叶植物多为厚壁组织。很多双子叶植物薄壁组织里面还有一圈多层厚壁组织鞘。到了小型叶脉通常厚壁组织消失,只剩下薄壁组织,成为维管束鞘。很多双子叶植物维管束鞘上下没有叶肉,但分布有没有叶绿体的薄壁组织,叫做维管束鞘延伸区。具有维管束鞘延伸区的叶脉分布较稀疏,因为它旁边的一部分表皮组织可以承担叶脉的功能。
- 单子叶植物的维管束鞘。如果一层那么是薄壁组织。如果两层那么外层薄壁组织内层厚壁组织。
- 小型叶脉的一个突出特点是含有薄壁组织【传递细胞】。在最小的叶脉中,木质部只剩下了管胞,韧皮部只剩下一个薄壁细胞。木质部的导管和薄壁细胞和韧皮部的筛管消失。最小叶脉的脉梢只剩下木质部,韧皮部也消失。
- 禾本科植物的叶。大多没有叶柄,由叶片和叶鞘构成。表皮经过硅质化,分为长细胞和短细胞。叶片的上表皮具有泡状细胞,在横切面上呈现锯齿状。
- 250
植物系统学
绿色植物系统发育综述
细胞内共生事件
- 共生——蓝细菌。蓝细菌没有有性生殖。共生使得质体基因组大大缩减。
- 只有一次最初的内共生事件。这次内共生事件导致了 原初植物 的产生。原初植物包括绿色植物、红藻、灰色植物。在灰色植物中,质体仍然有细胞壁。
- 第二次内共生和第三次内共生产生了 异鞭毛类。
各种各样的藻类
-
世代交替——配子体和孢子体。
-
红藻。具有藻胆素【这种色素使得它们可以在深水中光合作用】。大多数红藻是丝状体,通过穿孔而不是胞间连丝联系。任何时候,红藻都没有可以移动的细胞,因此它们表现出超复杂的生活史,可能具有两个形态和生态显著不同的二倍体阶段【果孢子体】。红藻大多数是配子体占优势的种类。 然而,它们的生命周期通常涉及三个阶段,其中一个额外的二倍体阶段:果孢子体。 Polysiphonia是Rhodophyta的模式生物1。果孢子体是红藻生命周期中的一个额外的二倍体阶段。 它最终会产生并释放二倍体果孢子,这些果孢子会发展成四分孢子体。 四分孢子体的某些细胞经历减数分裂产生四分孢子,然后循环重复1。
-
Chromalveolate。Alveolates泡膜虫。 dinoflagellates涡鞭毛虫。 Apicomplexa顶复合体虫。 Ciliata纤毛虫。 Foraminifera有孔虫。Stramenopiles异鞭毛类。
-
Chromalveolata是一个由Thomas Cavalier-Smith提出的真核生物超级群,它是对他在1981年首次提出的Chromista王国的改进。 Chromalveolata的起源被认为是一个双鞭毛(具有两个鞭毛的细胞)和红藻之间的二次共生。 这种结合导致了含有叶绿素c的质体1。
Chromalveolata包括非常重要的光合生物,如硅藻,褐藻以及动植物中的重要疾病病原体。 Chromalveolata可以分为泡膜虫和异鞭毛类23。
-
Stramenopiles(也称为Heterokonts)是一类生物,其特征在于具有硬质三部分外部毛发。 在大多数物种中,毛发附着在鞭毛上,在某些物种中,它们附着在细胞表面的其他区域,在某些物种中,它们已经次要丢失(在这种情况下,与Stramenopile祖先的相关性来自其他共享的细胞学特征或遗传相似性)1。
Stramenopiles是一个单系真核生物群,其特征在于具有两个鞭毛,其中未成熟的鞭毛具有三部分毛发。 该群包括21个类别,超过100,000种物种2。
Stramenopiles或异鞭毛类构成了原生生物中最多种类和多样性的谱系之一。Stramenopiles包括生态重要的藻类(如硅藻或大型多细胞褐藻),以及异养(例如bicosoecids,MAST组)和寄生(例如Blastocystis,卵菌)物种1。它们包括各种藻类原生生物,异养鞭毛虫,欧泊莱和密切相关的原生动物鞭毛虫(所有内生于其他生物中的);放射虫和卵菌2。
Stramenopiles代表SAR超级群中的三个主要类群之一,与Alveolata和Rhizaria一起。 Stramenopiles是真核生物;由于它们既不是真菌,动物,也不是植物,因此被归类为原生生物。 大多数Stramenopiles都是单细胞的,但有些是多细胞藻类,包括一些大型海藻,即褐藻。 该组包括各种藻类原生生物,异养鞭毛虫,欧泊莱和密切相关的原生动物鞭毛虫(所有内生于其他生物中的);放射虫和卵菌1。
-
褐藻属于异鞭毛类。具有呈现褐色的类胡萝卜素。所有褐藻都是多细胞的,并且都经历了世代交替:二倍体(2n)和单倍体(n)阶段交替。 单倍体和二倍体生活形态看起来非常不同,它们是异形的。 二倍体形式,即二倍体或孢子体,在这些生命周期中占主导地位1。墨角藻的单倍体阶段甚至完全消失,变成了配子减数分裂【像在动物中一样】。许多褐藻是丝状体,但也有的褐藻大且有很复杂的分化。
-
硅藻有很多化石种类,有很大的多样性。硅藻的细胞壁由两层精雕细刻的硅板构成。除了在一些雄性配子中,没有鞭毛。
-
Alveolates(意为“像蜂窝一样凹陷”)是一组原生生物,被认为是Eukarya内的一个主要支系和超级门。 它们目前与异鞭毛类和Rhizaria一起分组在具有管状嵴线粒体的原生生物中,该组被称为SAR1。Alveolates包括一些非常重要的光合生物,如渦鞭毛虫,褐藻以及动植物中的重要疾病病原体。 它们还包括一些非常熟悉且数量众多的原生生物群,包括纤毛虫,如草履虫和喇叭虫。 纤毛虫是异养原生生物中最多样的群体,拥有7000种物种。 另一个非常多样但不太熟悉的群体是有孔虫。 虽然普通人可能不熟悉这个群体,但它们对古生物学家来说非常重要,他们使用它们的化石来确定岩石的年代,确定古气候并寻找油气储藏。 第三组是渦鞭毛虫。 这些泡膜虫是光合作用的,能够从阳光,二氧化碳和足够的溶解营养中制造自己的食物。 渦鞭毛虫以其周期性人口爆发而闻名,这会导致“红潮”,可能会杀死鱼类和贝类。 Alveolates的最后一组是顶复合体虫,这是一组寄生性和致病性原生生物。 它们以具有单细胞生物中最复杂的生命周期而闻名1。
-
涡鞭毛虫具有两条鞭毛,位于埋在细胞壁中的纤维素板间的特有凹槽中。包括甲藻。虫黄藻和珊瑚共生,共生形态缺乏纤维素板。可以引发赤潮和水华。Dinoflagellates(希腊语δῖνος dinos“旋转”和拉丁语flagellum“鞭子,鞭打”)是单细胞真核生物的一个单系群,构成了Dinoflagellata门,并且通常被认为是藻类。 Dinoflagellates主要是海洋浮游生物,但它们在淡水栖息地中也很常见。 它们的种群随海表温度,盐度和深度而变化。 许多甲藻都能进行光合作用,但实际上这些中的很大一部分都是混合营养的,将光合作用与摄食猎物(吞噬和吸噬)相结合1。
绿色植物
- 绿色植物是单系的。共衍征包括藻胆素的丢失、叶绿素b的合成、淀粉粒储存糖类,运动细胞具有多为两个的尾鞭型鞭毛并在基部有一个特征性的星形结构。
绿藻
- 绿藻包括绿藻纲,石莼纲,和Trebouxiophyceae。
- 绿藻纲包括一个团藻路线 volvocine 进化支,共衍征是超微结构。石莼纲共衍征是多核细胞的产生。Trebouxiophyceae一般缺乏运动细胞,许多种类适应陆生生活,和地衣或者无脊椎动物关联。
链型植物Streptopohytes
- 共衍征是纺锤丝指向成膜体。
植物生理学
植物细胞的亚显微结构和功能
- 细胞壁包括半纤维素交联的纤维素网络;果胶基质;伸展蛋白【伸展蛋白不是扩展蛋白】。半纤维素是多糖的混合物,包括木聚糖。果胶主要是alpha 1,4糖苷键连接的半乳糖醛酸。
植物的水分生理
- 蒸腾速率【gH2O/(m2 h)】、蒸腾效率【每kg水】、蒸腾系数【每g干物质】。
- 气孔运动的五种假说。淀粉与糖积累假说,K+积累假说,苹果酸生成假说,玉米黄素假说,蔗糖调节气孔运动假说【气孔的张开和K+有关,而开放的维持和关闭和蔗糖有关】。蔗糖来源于叶绿体淀粉的水解或者质外体经过H+/蔗糖同向转运体
植物的矿质营养
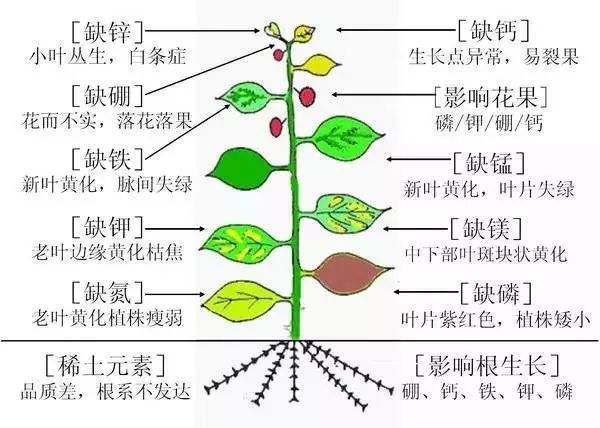
-
氮磷钾。氮肥:植物叶大绿,分支多,营养生长。磷肥:促进各种代谢。钾肥:品质元素。
-
钾肥是品质元素。缺少硼元素导致华而不实。
-
缺素症:
![作物缺素症状综合整理]()
缺锌植物不能合成色氨酸,缺少AUX。缺铜时,叶子黑绿色,并有坏死点。
植物的呼吸作用
- 乙醇酸氧化途径——GAOP是水稻等能适应淹水条件的植物根系特有的糖降解途径。关键酶是乙醇酸氧化酶。目的是氧化土壤中的还原性物质,防止毒害。通过乙酰CoA——乙酸——乙醇酸——乙醛酸——草酸——甲酸和CO2的氧化产生H2O2。氧化土壤。
- 呼吸链抑制物:复合物I——鱼藤酮、巴比妥酸。复合物II——TTFA、萎锈灵。复合物III【细胞色素c氧化还原酶】——抗霉素A【AA】。复合物IV——CO,CN-,N3-【通过和Fe结合】。
- 呼吸链电子传递。对鱼藤酮不敏感的NADH脱氢酶——氧化苹果酸穿梭的NADH,产生6H+,P/O=1.5。外源NADH电子传递途径——同上。交替途径——抗氰支路。交替氧化酶是唯一组分。复合体I传给UQ后直接传给AOX。P/O=1。线粒体外氧化酶——乙醇酸氧化酶、多酚氧化酶、抗坏血酸氧化酶。
- 末端氧化酶。细胞色素氧化酶——即复合体IV。含有铁卟啉和铜。交替氧化酶——抗氰氧化镁。位于线粒体内膜。含铁。二硫键非共价结合的还原型二聚体活性高。被水杨基氧肟酸抑制。酚氧化酶——质体和微体。含有铜。马铃薯、苹果变成褐色就是酚氧化酶的结果,和红茶制作有关。被CN-和CO抑制。抗坏血酸氧化酶——细胞质。乙醇酸氧化酶——黄素,不含有金属。被CN-抑制不被CO抑制。
- 呼吸跃变和乙烯、抗氰呼吸有关。呼吸跃变标志着果实进入衰老阶段。
- 水稻在开花初期,以EMP-TCA为主。随着籽粒成熟,PPP逐渐加强。
- 萜类化合物包括除虫菊、沉香醇、植醇、赤霉素、冷杉醇、番茄红素、棉酚、紫杉醇、杜仲胶、橡胶。酚类化合物包括咖啡酸、阿魏酸、绿原酸、水杨酸、植保素、香豆素、黄酮、黄酮醇、花色素苷、异类黄酮、木质素、鞣质和醌类。含氮次级化合物包括吗啡、奎宁、咖啡因、烟碱、可卡因、可可碱、秋水仙碱、生氰苷、非蛋白质氨基酸。
- 块茎、块根在贮藏期间处于休眠状态,而不是像果实那样处于成熟状态。
- 夏季早稻灌浆期灌注跑马水降温,可以降低呼吸消耗,有利于种子成熟。
植物的信号转导
生长素
- AUX包括IAA,2,4-D,PAA。
- IAA在分生组织和幼嫩的分裂组织中合成。检测IAA存在的方法是将beta-葡萄糖醛酸酶 GUS报告基因和一个对生长素有反应的启动子后面,显现蓝色。
- IAA的生物合成就是要把色氨酸变为吲哚乙酸。也就是要把丙氨酸变为乙酸。主要通过吲哚丙酮酸途径。还有色胺途径、吲哚乙醛肟途径、吲哚乙酰胺途径。
- IAA通过共价结合形成结合态IAA,包括肌醇、葡萄糖、氨基酸、多肽、糖苷、糖蛋白等。
- IAA的降解包括酶氧化降解和光氧化降解。酶氧化降解的脱羧降解需要Mn++和一元酚。光氧化降解需要核黄素。
- IAA运输的主要部位是木质部。一个例外是胚芽鞘通过非维管束途径运输。用化学渗透模型解释。
- 化学渗透模型。简要地说,IAA的吸收和外流的载体蛋白具有极性。IAA的吸收依赖跨膜的质子梯度,IAA的流出依赖跨膜电位【一旦进入细胞,IAA就变成阴离子,只可能通过主动运输流出细胞】。AUX/LAX蛋白参与IAA的吸收。PIN蛋白参与IAA的流出。它们分布具有极性。而ABCB参与生长素的流出,但不具有极性,均匀分布在细胞上。ABCB的作用是为了抵抗IAA流回细胞,维持IAA流。
- IAA参与了茎、根的伸长生长。维管系统的分化——高浓度促进木质部,低浓度促进韧皮部。植物的向性——向光性和负向重力性。顶端优势的维持。抑制花的形成,增加雌花,单性结实。
- 酸生长理论。IAA激活质子泵,激活扩展蛋白。扩展蛋白使得细胞壁松弛。
- 基因激活假说。细胞壁成分的合成。生长素结合蛋白ABP1主要位于内质网,但不重要。在核内,ARF是转录因子,和IAA反应元件结合。AUX1/IAA和ARF结合抑制活性。在IAA存在时,IAA作为分子胶水把AUX1/IAA和SCF-TIR1/AFB【E3】结合,导致AUX1/LAX降解。从而ARF和IAA反应元件结合。
赤霉素
- 赤霉素是一种双萜。GA有很多种,但只有很少一部分具有生理活性。GA第7位碳原子上的羧基是GA所共有的,也是发挥功能所必需的。
- GA的合成部位有生长中的果实和种子,幼茎顶端和根部。
- 所有的GA合成都必须经过一个共同前体——GA12-7-醛。经过环化。矮壮素CCC、缩节胺可以抑制环化步骤。
- 结合赤霉素可以和糖结合。酸水解和酶分解可以释放出自由GA。
- GA主要通过韧皮部运输。
- AUX只能促进离体组织的伸长生长,GA能促进完整植株的伸长生长,使得节间伸长。种子萌发需要GA激活alpha-淀粉酶。GA促进雄花分化。
- 在核内,DELLA抑制GA响应。GA和受体GID1结合后促进它泛素化DELLA,使得GA响应。
植物的生长生理
- 植物激素对细胞生长的作用:CTK——细胞横向生长。IAA,GA——细胞伸长。ABA,ETH——抑制细胞伸长。
- 植物激素对细胞分化的作用:CTK/IAA低时生根,高时生芽,中等时愈伤组织。AUX和CTK诱导愈伤组织分化为木质部。
- 蔗糖使韧皮部分化,低蔗糖使得木质部分化。
- TTC法可以测定种子的生活力。活种子被染成红色。
- 种子活力是指在非理想条件下,迅速、整齐产生健壮的植株的能力。种子活力和种子生活力是不一样的。
- 干燥种子最初的吸水通过吸胀作用。蛋白质、淀粉和纤维素吸水的能力依次递减。
- 需光种子如杂草蔬菜。需暗种子如茄科、葫芦科。需光种子被红光促进萌发。
- 种子萌发的吸水包括三个阶段。一阶段吸胀作用,快。二阶段慢,呼吸也停滞在一个水平,但种子在二阶段的代谢活动非常旺盛。三阶段快,因为胚根突破种皮。
- 干燥种子中的酶原经过水合被激活,而不是水解。【水解激活并不普遍发生在植物细胞,而是动物细胞。】编码早期蛋白质的mRNA显然被贮藏在细胞质中。被称为长命mRNA,可以和蛋白质结合成为信息体。
- 糊粉层中淀粉酶的水解通过GA激活。
- 植酸是六磷酸肌醇。是磷元素的主要贮存形式,也是其他多种矿质元素的贮存形式。
- 对种子进行渗透处理需要用PEG【聚乙二醇】。
- 在夏季植物晚上生长快,在冬季植物白天生长快。
- ABA和IAA沿着维管束上行。
- 顶端优势。营养学说和生长素学说。IAA,GA促进顶端优势,CTK抑制。在根部的顶端优势CTK促进,ABA抑制。
- 多次开花植物具有大小年现象。
- 营养生长用氮肥,生殖生长用磷肥、钾肥。
- 植物生长的最适温度并不是植物生长最健壮的温度。而是协调最适温度,比生长最适温度稍低。
- 在微重力环境中,植物的茎基本没有,根乱转。
- 光敏色素包括黄化组织的PI和绿色组织的PII。是易溶于水的浅蓝色色素蛋白。具有生色团,是四个线性排列的吡咯环。硫酯键相连。生色团在黑暗条件下,先在质体中合成,然后在细胞质基质中和脱辅基蛋白结合。Pr和Pfr之间的转换包括光化学反应和黑暗反应。黑暗反应需要水,这就是为什么干燥种子没有光敏色素反应,但是被水浸泡过的种子却具有光敏色素反应。
- 光敏色素反应包括极低辐照度反应VLFR【拟南芥种子的萌发】,低辐照度反应LFR【莴苣种子的萌发、叶子的运动】,高辐照度反应HIR【黄化苗的绿化,下胚轴的伸长,花色素的形成】。VLFR和HIR不可被远红光逆转。LFR可以。、
- 快反应包括Pr和Pfr的分布和转板藻叶绿体转动。
- 红光导致自由IAA减少,Eth合成减少,CTK合成增加。远红光促进Eth合成。
- 光敏色素作用的膜假说【小叶运动依赖质子泵、叶绿体运动依赖钙离子】和基因调节假说【光敏色素是一个自催化的丝苏氨酸激酶】。
- 蓝光受体具有三指状作用光谱。包括隐花色素、向光素、玉米黄素。隐花色素可以介导去黄化、光周期、生物钟、基因表达,生色团为FAD和蝶呤。向光素介导植物体的运动,生色团为FMN。玉米黄素参与了气孔开闭的调节。
- 光周期和生物钟通过光敏色素和隐花色素共同调控。
- 紫外光B受体通过刺激黄酮和花青素形成介导植物的在辐射下的自我保护。
- 向光运动由向光素介导,蓝光诱发。通过形成电势差,把带有负电荷的IAA吸引到背光面。向光素也是丝苏氨酸激酶。隐花色素的作用类似胆固醇对流动性。
- 横向光性通过K+控制叶枕的运动细胞引起。向日葵的向阳运动远比向光运动复杂。
- 向重力性由IAA,Ca+和ABA介导。IAA由电势差聚集到下侧,抑制下侧生长。平衡石通过刺激内质网,将钙离子释放到细胞质,引发下侧的IAA积累。ABA也在下侧积累。
- 感夜性和感热性是生长性运动。感震性则是膨胀性运动。
植物的生殖生理
- 成花过程可以分为成花诱导、成花启动、花的发育三个阶段。
- 植物开花之前必须达到的状态称为花熟状态、与之对应的是幼年期。幼年期生长快、呼吸旺盛、核酸代谢和蛋白质合成快。花芽分化就是营养生长转向生殖生长的标志。
- 春化作用指的是幼苗或者种子。感受部位在茎尖生长点。需要春化的植物包括大多数二年生植物。春化中途高温导致去春化作用。春化需要氧气、水分和糖类。在春化前期细胞色素氧化酶为主,后来变为多酚氧化酶和抗坏血酸氧化酶。春化作用的分子机制。最重要的效应蛋白是FLC。FLC抑制开花。VIN3抑制FLC。低温春化、VRN通过促进FLC去抑制开花【促进途径】。低温春化还可以通过抑制FRI去抑制开花。【抑制途径】
- 花诱导和花形成需要的日照强度不同,称为双重日长。
- 临界日长实际上是临界暗期。长日植物实际上是短夜植物,短日植物实际上是长夜植物。虽然暗期起到决定性作用,但是光期也不可缺少。
- 光敏色素是光周期调控决定性色素。红光最有效,可以被远红光逆转。蓝光作用小。感受光周期的部位是叶片。
- 成花素的本质是GA和开花素【FT蛋白】。不同光周期反应的植物成花素都一样。GA是长日植物开花的限制因子,开花素是短日植物开花的限制因子。开花最重要的步骤是FT蛋白到茎顶端分生组织结合转录因子FD,形成FD/FT复合物,诱导开花。
- IAA抑制短日植物开花,促进长日植物开花。CTK都促进。ABA促进短日植物开花,抑制长日植物开花。BR使得未春化的植物花期提前。C/N比高的植物开花。
- 成花诱导的途径包括 GA途径、自主途径、春化途径、光周期途径。自主途径和春化途径最重要的蛋白是FLC。FLC抑制开花。光周期途径通过CO,FT,FD促进开花。
- 硼元素对花粉萌发和花粉管生长有重要作用。
植物的逆境生理
- 胁迫因子首先使得生物膜受害。包括原处直接伤害【质膜透性增大,细胞脱水】、原处间接伤害【酶代谢紊乱】、次生胁迫伤害【如盐害中的水分胁迫】。
- 相容性物质不干扰细胞内的正常生化反应,参与渗透调节。包括脯氨酸、甜菜碱、蔗糖、山梨醇、甘露醇、多胺。其中,脯氨酸还可以作为氮源、氮汇;也可以增强蛋白质的水合作用,减少蛋白质的沉淀。甜菜碱是一类季铵碱。
- 逆境下,ABA和乙烯上调。乙烯可以提高多酚氧化酶、几丁质酶活性;引发器官衰老和枝叶脱落。
- 冷害引起植物呼吸速率忽高忽低。
- 缓慢降温时,膜均一凝固和紧缩,膜透性减小,吸水困难,造成生理性干旱。突然降温时,膜不均一凝固和紧缩,导致膜的破裂,透性增加,导致细胞内容物外渗。ABA,CTK,2,4-D,油菜素内酯可以提高抗冷性。施加P,K,少施加N可以增加抗冷性。
- 冻害发生时,最先受到伤害的是叶绿体膜。然后是液泡膜。最后是质膜和线粒体膜。冻害也会导致蛋白质不正确二硫键的形成。植物组织抗冻性的基础在于组织蛋白质分子间二硫键形成。生长延缓剂可以提高抗冻性。
- 冷驯化包括寒驯化和冻驯化。
- 有机酸能够抗热,通过减轻氮的毒害。油料种子的抗热性高于淀粉种子。
- 涝害时,乳酸积累是导致细胞酸中毒的主要原因。细胞将会抑制乳酸脱氢酶、促进丙酮酸脱氢酶。促进将丙酮酸转变为乙醇。
- 根据抗盐性可以把植物分为盐生植物、甜土植物、真盐生植物。IAA可以提高植物的抗盐性。
- 植物病害可以发生超敏反应。超敏反应是一种细胞程序性死亡。植物对病害的抑制物质包括植保素、木质素、抗病蛋白、几丁质酶、苯丙氨酸解氨酶、beta-1,3-葡聚糖酶、植物凝集素等
- 系统获得性抗性是指植物一部分被侵染后,全身的抗性都增强的现象。需要R基因,对应的病原菌分子则被称为激发子。虫害对应的有分子系统素。
- 光化学烟雾包括O3,NO2,RCHO,PAN。
动物学
总论
-
卵裂类型:【具体图示见发育生物学P12 】
- 完全卵裂
- 少黄卵——辐射卵裂。棘皮动物、头索动物、半索动物。
- 中黄卵——
- 辐射卵裂——两栖类、圆口类、软骨硬鳞鱼【鲟鱼】。
- 螺旋卵裂——软体动物【除了头足类】、环节动物【除了多毛类】、海绵动物、扁形动物、毛颚动物、假体腔动物、【对虾】。
- 两侧对称——腔肠动物、尾索动物
- 转动型——哺乳动物、线虫。【胚胎致密化。另外一个需要注意的是,因为其独特的排列方式,导致转动卵裂完成的囊胚长得很像中黄卵。】
- 不完全卵裂
- 盘状卵裂——端黄卵——鱼类、头足类【双边不等裂】、爬行类、鸟类、卵生哺乳类、蝎目、【卵齿蟾】。
- 表面卵裂——中央黄卵——节肢动物【除了肢口纲】【早期为合胞体囊胚】
- 完全均等卵裂——钵水母类、腹足类、肢口纲、棘皮动物、半索动物、头索动物、毛颚动物、链蝎。
- 多毛类——端黄卵,螺旋卵裂。
- 完全卵裂
原生动物门
-
- 寄主:人和白蛉子
- 白蛉子内:
- 在消化道内有许多前鞭毛体
- 人内:
- 在人体内脏的巨噬细胞内,有许多无鞭毛体。
-
- 寄生阶段——大滋养体、小滋养体;寄生在肠腔。
- 感染阶段——4个核的包囊
- 消灭包囊和防止包囊进入体内是防治的根本环节
-
- 具有石灰质或其他物质形成的外壳
- 有世代交替
- 现在的海底大约有 35% 的泥沙来自于有孔虫的外壳沉积,放射虫也是如此。
-
- 寄主:人和按蚊
- 按蚊内:
- 胃腔:大小配子母细胞形成配子;受精产生合子
- 胃壁基膜和上皮细胞之间:动合子发育为卵囊,内含卵囊
- 人体内:在肝细胞和红细胞中发育
- 红细胞前期:子包子 - 滋养体 - 裂殖子——当裂殖子成熟后,破裂肝细胞而出,才能侵入红细胞。
- 红细胞外期——疟疾复发的根源
- 红细胞内期:环状体 - 大滋养体 - 裂殖体;以红细胞内的血红素为养料
海绵动物门
- 毛壶卵裂属于不等裂。
线虫动物门
环节动物门
-
口前叶和尾节不属于体节。节生长区位于尾节前的区域。身体生长是从后端连续增加体节的结果。
-
系膜的结构类似于隔膜,是每一体节左右体腔膜接触形成的。
-
体腔细胞有防卫功能,气体运输功能。
-
疣足是体壁向外突出的扁平叶状结构,体腔也伸入其中。
-
刚毛是由几丁质形成的chaeta, 和节肢动物的 seta 不同:
seta chaeta 内有中空小管 内有刚毛髓腔 β - 几丁质 α - 几丁质 连有成刚毛细胞 连有感觉细胞 有刚毛缩肌 没有刚毛缩肌 机械运动功能 机械感觉功能 -
有的环节动物具有原肾管,其原肾管不具有焰细胞而具有管细胞。如多毛类的原肾管和体腔管结合。
-
环节动物的中枢神经系统
- 脑神经节或咽上神经节。
- 前脑:口前叶感受器
- 中脑:眼、口前叶触手、吻、咽
- 后脑:项器
- 围咽神经:前脑和中脑发出
- 咽下神经节:身体运动控制中心。调节远距离体节运动,启动运动。
- 脑神经节或咽上神经节。
-
副神经索中存在巨纤维。巨纤维的功能只在于逃避反应,而不是正常的运动。
-
沙蚕口前叶后端两侧有一对纤毛窝,称为项器,是化学感受器,有嗅觉功能。
-
沙蚕体壁每节都有两束连结正腹部和背侧部的背腹斜肌,牵动疣足运动。
-
血液在 背 血管中由后往 前,在 腹 血管中由前往 后 流动。
-
沙蚕有特殊的生殖现象——生殖态(身体后部变为生殖节)和群游现象(同时从海底游向海面)。
-
螺旋虫属于管栖触须超目
-
蚯蚓定位:【受精贮卵雌环雄】
-
蚯蚓的背腹肠系膜退化。
-
蚯蚓的消化系统:
-
咽部有单细胞咽腺分泌粘液和蛋白酶
-
食道 壁有钙腺,司离子调节,也可以中和食道的酸性物质
-
有嗉囊和砂囊,嗉囊薄壁,砂囊厚壁。
-
肠盲道可以增加消化面积
-
锥形盲囊能分泌多种消化酶,是重要的消化腺。从第26或者27体节分出。
-
黄色细胞——来源于体腔膜细胞。有类似肝的作用。
-
隔膜小肾管、咽头小肾管、体壁小肾管:
-
隔膜小肾管——有肾口,后部体节,肾孔开口于肠内。
-
无肾口——
- 咽头小肾管:前部,肾孔开口于咽。
- 体壁小肾管:广泛分布,肾孔开口于体表。
-
蛭纲的嗉囊就是胃,发达,用来存储血液。
-
蛭纲消化管分泌物大多没有消化酶,和嗜水气单胞菌共生。
-
蛭纲的前后端神经节集中愈合,在前部产生咽上神经节和咽下神经节;在后部产生尾神经节。
软体动物门
节肢动物门
-
蜘蛛的马氏管是内胚层,昆虫的马氏管是外胚层来源的。
-
节肢动物的体壁包括基膜、单层上皮细胞、表皮。表皮包括:
- 上表皮,含脂蛋白,不含几丁质,防止水分丧失或进入体内。
- 原表皮:
- 外表皮:蛋白质鞣化。
- 内表皮:柔软。
-
附肢。包括原肢和端肢。
- 双枝型——还有内叶和外叶。
- 单枝型——没有内叶和外叶。
-
鳃和书鳃——鳃是体壁向外凸出产生的,书鳃是体壁向内折叠产生的。
-
血液通过心孔回心。
-
昆虫没有呼吸色素。
-
甲壳纲
- 额剑——体壁突起,不是附肢。交接器——雄虾第一腹肢的内肢特化
- 第一触角——触觉、嗅觉、平衡。第 2 小颚——呼吸。
- 胃磨,肝胰脏。
- 触角腺(绿腺)。腺体部分的内端是一个盲囊——端囊,代表残留的体腔。有排泄和回收离子功能。但蛋白质代谢产物氨通过鳃排出。幼体有小颚腺。
- 受精囊——4,5。雌性生殖孔——3。精巢开口——5。
- 内分泌:
- 蜕皮:小颚的 Y器 分泌 蜕皮激素MH 使得动物蜕皮。眼柄中的 X器 分泌 抑制蜕皮的激素MIH 抑制动物蜕皮,储存到窦腺中。
- 体色:色素颗粒集中体色变深。来源于胡萝卜素衍生物,和蛋白质结合成为虾青素。
- 生殖:
- 雌性:窦腺——抑卵巢激素GIH。中枢神经系统——促卵巢激素GSH。
- 雄性:输精管末端,促雄腺。被 X器 抑制。
-
鳃足纲多淡水生活,胸足可以调节渗透压。蚤状溞孤雌生殖。藤壶雌雄同体。
-
软甲纲鼠妇具有伪气管。
-
肢口纲有唇状瓣、生殖厣,是附肢。有尾剑,不是附肢。
-
蛛形纲有吸胃和中肠盲囊。马氏管来自于内胚层。氮代谢产物鸟嘌呤和尿酸。蝎子有栉状板,感觉。蛛形纲有生殖蓋板,纺绩突,纺绩器——为附肢。棉红蜘蛛是螨。
-
多足亚门有唇基和上唇,但不是附肢。没有背甲,上皮层缺少脂质和蜡质,保水能力差。消化管长而直,没有盲囊。唇足纲生殖孔位于倒数第二节。
-
六足亚门
-
有上唇和舌,不是附肢。上颚、下颚、下唇是附肢。雄性外生殖器是 9体节附肢。雌性产卵瓣是 8,9 体节的附肢。
-
鼓膜听器:蝗虫和蝉——腹部;蟋蟀——前足胫节;蛾类——胸部和腹部交界。
-
昆虫的光感受器:
-
唾液腺。
-
气管——螺旋丝,端细胞,微气管有液体。
-
血细胞有吞噬功能。心脏搏动能力不强,需要依靠身体活动。
-
直肠腺。
-
精荚。
-
马氏管来自于外胚层。
-
昆虫的口器:
-
虹吸式——下颚
-
舐吸式——下唇
-
刺吸式——上下颚特化为细长的口针,上颚包裹下颚。下唇也加长,有凹槽容纳上下颚。
-
嚼吸式——上颚。
-
昆虫内分泌:
-
神经分泌细胞——促蜕皮激素【脑激素、活化激素】。心侧体储存。
-
咽侧体——保幼激素。
-
前胸腺——蜕皮激素。
-
半翅目蚜虫孤雌生殖,胎生。介壳虫固着生活。
触手冠动物
棘皮动物门
- 棘皮动物的体腔:左右体腔囊分为前体腔囊、中体腔囊、后体腔囊。左前体腔囊——轴窦,左中体腔囊——水管系统,后体腔囊——体腔。
- 水管系统——筛板、石板、环管、侧管、辐管、管足、
- 血系统——环血管、辐血管、轴线、反口环血管
- 围血系统——环窦、辐窦、轴窦
- 神经系统包括口神经系、下神经系、反口神经系。其中,下神经系和反口神经系由中胚层发育,司运动。口神经系来源于外胚层,司感觉。
- 皮鳃和体腔相同,有呼吸和排泄的功能。
- 海盘车具有幽门盲囊,消化主要在幽门胃进行。海盘车肛门已经没有排遗功能,不能消化的食物从口吐出。
- 内陷法。
- 海胆纲具有口管足。亚里士多德提灯。
- 蛇尾纲消化道退化,食道短,后连有盲囊状的胃。没有肠【海盘车有肠】。没有肛门。
- 海参纲具有疣足。有胡希恕和居维尔氏器。水管系统中流动体腔液。【其他棘皮动物水管系统中流的是海水】
- 毛颚动物半透明,没有循环系统和排泄系统。是唯一具有复层上皮的无脊椎动物。雌雄同体。
- 棘皮动物的幼虫:海星——羽腕幼虫、海蛇尾和海胆——长腕幼虫、海参——耳状幼虫、海百合——樽形幼虫。
半索动物门
- 半索动物门全部海产。
- 体腔分化为吻腔、领腔、躯干体腔。
- 具有口索,并不是退化的脊索。
- 没有胃的分化。
- 开管式血液循环。血管小球突入吻腔中,是排泄器官。
- 背神经索在渗入领的部分出现狭窄空腔,曾经被认为是背神经管的雏形。
- 具有柱头幼虫,和海参的短腕幼虫很相似。
脊索动物门
- 脊索在发生上来自于胚胎的原肠背壁。脊索细胞具有大液泡,缺少细胞间隙,类似于植物细胞。
- 尾索动物亚门具有外套膜和被囊,被囊素类似于植物纤维素。整个动物界中具有被囊的动物只有尾索动物和少数原生动物。
- 围鳃腔的内壁是外胚层。
- 具有背板【即咽上沟】、内柱。
- 开管式循环系统。没有动静脉之分,血液双向流动。这种血液循环方式是绝无仅有的。
- 雌雄同体。
- 神经中枢是一个神经节。
- 逆行变态。
- 头索动物亚门具有口笠,口笠内的空腔叫做前庭。前庭内壁具有轮器。搅动水流进入口内。
- 具有肝盲囊,可能和肝是同源器官。肠内具有回环结,在这里搅动食物。内助细胞富集碘,和甲状腺同源。
- 没有心脏。腹大动脉和鳃心具有搏动功能。
- 排泄器官是按体节排列的肾管。此外在咽部后端背侧各有一个褐色漏斗体。可能具有排泄的功能。
- 神经系统包括脑泡和 2 条脑神经。
- 神经管两侧的脑眼是光线感受器。神经管前端,单个的眼点没有视觉作用。可能是退化的平衡器官或者遮光物。
- 哈氏窝和脑下垂体同源。产生促性腺激素释放激素。
- 神经管刚产生时和原肠相通,为神经肠管。后来神经肠管闭塞。
- 围鳃腔的形成:身体腹面的一对纵行腹褶延伸相连,中间形成一个腔隙【体外】,前端关闭,后端形成腹孔,是围鳃腔唯一的开口。
圆口纲
- 头顶部具有单个鼻孔。
- 七鳃鳗具有口漏斗。
- 终身保留脊索,有两对软骨椎弓
- 没有完整的软骨脑颅
- 嗅囊软骨和听囊软骨独立。
- 具有软骨鳃篮。在末端具有保护心脏的杯状围心软骨。不分节。4+9结构。
- 呼吸管入口具有 缘膜。没有胃 的分化。肠管内有螺旋状的黏膜褶。具有 肝。成体 没有胆囊。没有独立的胰脏,胰细胞散在。
- 呼吸管有内鳃孔,鳃囊,外鳃囊。鳃囊来自于内胚层【】。盲鳗外鳃孔先通入总鳃管。
- 七鳃鳗成体附着在鱼身上时,水流的进出都通过外鳃孔。不附着在鱼身上则从口进入,外鳃孔出。
- 出现心脏。
- 脑排列在一个平面上。脑神经10对,但不具有视交叉。
- 只具有内耳。七鳃鳗具有两个半规管。盲鳗具有一个半规管。
- 瞳孔大小不能调节。晶体适合看近物。视觉调节通过角膜肌看远物,将角膜拉平。松果眼位于间脑顶部。
- 七鳃鳗雌雄异体,没有生殖导管。盲鳗雌雄同体,成体退化其中之一。
- 盲鳗只有一个半规管,没有呼吸管的分化,鳃篮退化,成体前肾,雌雄同体,没有变态。七鳃鳗有两个半规管,成体后位肾,有变态。
鱼纲
- 鱼类离水后会因为鳃的粘连和干燥,窒息死亡。
- 软骨鱼类眼后有 喷水孔,为第一对鳃裂。有泄殖腔。硬骨鱼有泄殖孔和肛门。雄鲨的腹鳍内侧具有鳍脚,是交配器官。
- 多细胞腺有毒腺和发光器。
- 软骨鱼的肩带不和头骨或者脊柱直接关节,称为 肩胛乌喙骨。
- 鲤形目最后一对鳃弓为下咽骨,具有 咽齿,和基枕骨腹面的角质垫研磨食物。
- 多数硬骨鱼类偶鳍骨中的基鳍骨消失,辐鳍骨退化或不存在,真皮鳍条多直接连接在带骨上。
- 眼球肌——外展神经,外直肌。滑车神经,上斜肌。动眼神经,上直肌,下直肌,下斜肌,内直肌。
- 发电器官:电鳗——轴下肌。电鲇——皮肤腺。
- 软骨鱼具有螺旋瓣。有肝和胰脏。肝大,调节身体密度。
- 硬骨鱼类具有鳃耙。肉食性胃发达并具有幽门盲囊【如鲈形目,“桂花”】。鲤科没有胃,肝和胰脏散在成肝胰脏。
- 鱼类主要通过鳃盖的运动完成呼吸。
- 硬骨鱼类有鳔。快速游泳的金枪鱼和底栖的鮟鱇没有鳔。位于肠管背侧。
- 闭鳔类通过卵圆区和红腺调节鳔的大小。红腺:腹腔肠系膜静脉通入,肝门静脉返回;卵圆区:背大动脉通入,后主静脉返回。
- 肺鱼的鳔静脉直接返回心脏的左侧,有心房和心室不完全间隔
- 鲨鱼在背大动脉两侧各有一个淋巴管。硬骨鱼类在最后一枚尾椎骨下方有两个相连的淋巴心,有脾脏。
- 软骨鱼类有有泄殖腔,没有膀胱。硬骨鱼类有肛门和泄殖孔,有膀胱。
- 软骨鱼类输卵管的前端具有壳腺。
- 角鲨卵胎生。星鲨假胎生【卵黄囊胎盘】。黄鳝、剑尾鱼性逆转。
- 软骨鱼类大脑嗅觉。中脑是视觉中枢和最高级中枢。小脑是运动调节中枢。延脑是多种生理功能和感觉(听觉、皮肤感觉、侧线感觉)、呼吸、色素细胞作用调节的中枢。延脑前部有耳状突,是听囊和侧线的感觉中枢。视交叉后方有下叶,下叶后有血管囊。是鱼类特有的压力感受器,可以感受水的深度。
- 软骨鱼类没有完整的交感神经干。硬骨鱼类具有。但硬骨鱼类的副交感神经原始,只随3,10对。软骨鱼类随3,6,9,10对脑神经。
- 软骨鱼类具有罗伦瓮,是电感受器官
- 鲨鱼的眼反光结构是亮层,位于脉络膜。适合看远物。视觉调节看近物。通过 晶状体牵引肌 调节 晶状体的前后位置。硬骨鱼类反光结构是银膜,位于脉络膜和巩膜之间。适合看近物。视觉调节看远物。通过 镰状突【硬骨鱼类特有,是脉络膜的一个膜质突起。前端伸出一个晶状体缩肌】 向后移动晶状体。
- 全头亚纲体表光滑无盾鳞。上颌和脑颅愈合。有泄殖孔和肛门。腔棘鱼鳔退化。肺鱼有内鼻孔。腔棘鱼和肺鱼脊索发达无椎体。
- 泥鳅、鲫鱼属于鲤形目。海马属于刺鱼目。黄鳝属于合鳃目。带鱼、银鲳、丽鱼、弹涂鱼、金枪鱼、黄鱼、翻车鱼属于鲈形目。鲈形目具有幽门盲囊。
- 鲨鱼具有4个全鳃,9个半鳃,有鳃孔。硬骨鱼有8个半鳃,并有鳃盖。
两栖类
- 两栖类开始腰带直接和脊柱连接,表皮角质化,具有颈椎和荐椎,椎弓出现关节突,舌颌骨变为耳柱骨。
- 粘液腺是多细胞腺体,属于泡状腺,下沉到真皮层内。
- 具有两个枕髁【侧枕骨】,和哺乳类相同。脑颅背侧膜骨只有额骨、顶骨【青蛙愈合成为额顶骨】、鳞骨。脑颅腹面膜骨只有副蝶骨。颌弓为自接式。出现了耳柱骨。
- 现代两栖类肩带主要包括肩胛骨、乌喙骨、前乌喙骨和锁骨。腰带具有骨盆。但蛙类不具有骨盆,髂骨极度前伸,左右耻骨坐骨联合。
- 青蛙具有桡尺骨、胫腓骨、腓跗骨。前肢4指。
- 弧胸型和固胸型的区别是前乌喙骨是否交叉。
- 四足动物具有肌肉质舌和唾液腺。蛙类在口咽腔两侧或者底部具有声囊,声囊是发生的共鸣器。
- 两栖类具有一对囊状的肺。无尾两栖类的呼吸主要通过咽式呼吸。
- 动脉圆锥内具有螺旋瓣。
- 第三对动脉弓变成颈动脉,第四对体动脉,第六对肺动脉。注意有尾两栖类具有动脉导管连接体动脉和肺动脉。有的还具有颈动脉导管连接颈动脉和体动脉。
- 两栖类的后肢血液需要经过肾门静脉或者肝门静脉才能返回心脏。
- 两栖类淋巴系统在皮下扩展成为淋巴腔隙,具有两对可以波动的淋巴心,不具有淋巴结。
- 蟾蜍等两栖类雄性具有比德器,相当于退化的卵巢,去势后可以发育为正常卵巢。生殖腺前方具有许多脂肪体提供能量。
- 青蛙已经具有了可动的下眼睑和泪腺,并具有半透明的瞬膜。具有中耳。高频繁殖通过鼓膜,低频警告通过前肢传导。出现了犁鼻器,开口于内鼻孔和口腔的连接处。无尾目和无足目犁鼻器和鼻囊分开。
- 蚓螈没有鼓膜,头骨硬骨多,双凹型,长肋骨,无胸骨。多真皮骨质小鳞。体内受精。
- 有尾目双凹型或者后凹型,有肋骨和胸骨,没有鼓室和鼓膜。大多体内受精。鲵体外受精。
爬行类
- 两个胚外的腔:羊膜腔和胚外体腔。同时消化管后端突起产生尿囊。
- 头部产生颞孔。蜥蜴和蛇类头骨膜颅后院骨块消失,方骨露出。某些毒蛇方骨还可以和颅底骨块形成可动关节,并左右下颌骨前端通过韧带连接。
- 蛇不具有胸骨。爬行类开始具有两块荐椎。
- 颞肌和咬肌。爬行动物开始出现盲肠,和消化植物纤维有关。
- 鳄鱼心房心室完全分隔,但在左右体动脉基部具有潘氏孔。
脊椎动物比较解剖
骨骼系统——头骨及内脏弓
-
脊椎动物头骨的基本结构
- 软骨脑颅
- 枕骨区:基枕骨、上枕骨、外枕骨【侧枕骨】。围成枕骨大孔。
- 耳骨区:前耳骨,后耳骨、上耳骨——常和上枕骨愈合。
- 蝶骨区
- 底墙——基蝶骨、前蝶骨
- 侧墙——翼蝶骨、眶蝶骨
- 筛骨区
- 膜颅
- 头骨顶部:
- 枕骨区:后顶骨、间顶骨、膜质上枕骨
- 眼眶后区:顶骨
- 眼眶区:额骨
- 头骨侧部:
- 颊区:鳞骨
- 眼眶的四周:后眶骨、后额骨、上眶骨、前额骨、泪骨
- 后侧方未标示的骨骼
- 鳃盖骨——硬骨鱼特有
- 颞骨——早期四足动物具有,位于顶骨和鳞骨之间。
- 腭罩:
- 中央:副蝶骨、犁骨
- 两侧:翼骨 【在低等脊椎动物分化为外翼骨、内翼骨】 、腭骨。
- 头骨顶部:
- 软骨脑颅
-
次生颌:上颌前颌骨上颌骨腭骨翼骨方骨、下颌关节骨隅骨齿骨。
-
脊椎动物内脏弓的基本结构:
- 内脏弓的基本结构——咽鳃骨、上鳃骨、角鳃软骨、下鳃软骨、基鳃软骨
- 颌弓:腭方软骨 为上鳃软骨特化,麦氏软骨 为角鳃软骨特化。腭方软骨在前方骨化为腭骨、中间骨化为翼骨、后部骨化为方骨。因此有的人也叫做腭翼方软骨。
- 舌弓:舌颌骨为上鳃软骨特化,角舌骨为角鳃软骨特化,基舌骨为基鳃软骨特化。
- 鳃弓。
-
颌的悬挂类型:鱼类要求上颌有大的活动性;呼吸空气的动物需要咬合力的提高。
- 双联型——腭方软骨通过基突、耳突、筛突以韧带和软骨脑颅联合;同时通过舌颌骨和耳囊结合。现存的某些鲨鱼和化石、棘鱼类、古总鳍鱼类。
- 舌联型——高等软骨鱼类、硬骨鱼类
- 自联型——腭方软骨通过基突 [通过蝶骨区底部的突起——基翼突] 、耳突、筛突以韧带和软骨脑颅联合;肺鱼和四足动物。从双联型演化而来。
- 全联型——全头类。
-
头骨和内脏弓的发生:
- 早期——索旁软骨、索前软骨、极软骨、视囊、耳囊、鼻囊,枕软骨。
- 随后——
- 索旁软骨联合为基板。
- 顶索软骨棒出现,后方留有颅底窗。
- 索前软骨联合为筛板。前端产生鼻间隔。后端留有脑下垂体窗,内颈动脉孔穿过——在软骨鱼类、肺鱼、两栖类和哺乳类后来封闭;在其他类群开放,由膜颅的副蝶骨在腹面封闭。
- 眶软骨发育为脑匣侧壁:
- 通过耳前小柱【形成侧蝶骨】、眶后小柱、框前小柱 和 软骨底墙连结。
- 颅底的类型:
- 平底型:有宽阔的脑腔,向前直达鼻囊。软骨鱼类、原始辐鳍亚纲、肺鱼、两栖类。
- 脊底型:左右索前软骨联合构成 眶间隔,和鼻间隔连续。真骨鱼类、高等爬行类、鸟类。
- 中间类型:原始两栖类、真口鱼类;哺乳类眶间隔不发达。
- 顶墙:大多数脊椎动物只在枕骨区具有顶墙——由枕弓构成,完整顶盖,有枕骨大孔。软骨鱼类特化为软骨,顶墙完整。
-
内脏弓的发生:脏壁中胚层、神经嵴外胚层。
- 颌弓:
- 腭方软骨后端形成方骨、前端在原始硬骨鱼类中形成翼骨;在与基突相对的地方有 升突,形成上翼骨,在哺乳类中构成侧墙,称为翼蝶骨——和硬骨鱼脑颅侧墙的翼蝶骨没有同源关系。
- 麦氏软骨后方形成关节骨,前方在两栖类形成颐骨。
- 颌弓:
-
圆口类的头骨:
- 不具膜骨
- 单个鼻囊
- 口漏斗结构——有环状软骨【脑匣前方】,前背软骨、后背软骨【背后方】,唇软骨【环状软骨两侧】。
- 挫食动作——舌软骨
- 不分节的鳃笼
-
软骨鱼类的头骨:
- 全部由软骨构成,大部分具有顶墙,只在前方有前囱。
- 枕骨区——单个枕髁。
- 筛骨区——筛骨区前端有吻骨。
- 鳃弓一般为 5 对。
-
硬骨鱼类的头骨:
- 软骨脑颅
- 进化趋势:
- 骨化——许多骨化中心。【例外:肺鱼和软骨硬鳞鱼 次生软骨化】
- 辐鳍鱼类——骨块分离,眶间隔退化为隔膜,有一个大空洞——动眼肌室【位于脑匣底部的脑下垂体窗,在眶颞区和耳囊区之间】。
- 总鳍鱼:成体脑匣分为完全不同的前、后两部分,两者之间有可动的关节。
- 枕骨区——基枕骨,单个枕髁。上枕骨、基枕骨不发达。
- 孔——迷走神经和副神经 侧枕骨;听神经——耳囊内侧;颜面神经、三叉神经——前耳骨前端;后颞孔——原始硬骨鱼类和原始四足动物。
- 进化趋势:
- 顶罩
- 原始硬骨鱼类
- 十分完整,只有外鼻孔、眼眶、松果孔
- 后方横行软骨——后来退化;单个的膜质上枕骨,成对的后顶骨、案骨。
- 单个膜骨——后来退化;膜质筛骨、间鼻骨、间额骨。
- 侧线管穿行
- 膜质的鳃盖骨
- 头骨底部具有咽骨。
- 总鳍鱼
- 背中部:成对的鼻骨、腭骨、顶骨、后顶骨
- 围眶骨:前额骨、后额骨、后眶骨、轭骨、泪骨。
- 上颌缘:前颌骨、上颌骨、方轭骨。
- 轭骨参与围眶骨而不是上颌缘。
- 颊部有丰富的鳞骨——和辐鳍鱼不同。
- 肺鱼
- 单个膜骨发达
- 软骨化
- 辐鳍鱼
- 眶后的颊部不发达,没有鳞骨。
- 不具顶骨孔。
- 眼眶大,眶前区域小
- 颌关节趋向前移、口裂缩小
- 口缘齿消失
- 颌骨有时不参与口缘。
- 原始硬骨鱼类
- 腭罩
- 古总鳍鱼
- 副蝶骨——脑匣腹部中央。副蝶骨后缘——在此处脑匣前半部分和后半部分关节。顶骨和后顶骨也有关节。
- 翼骨发达。
- 犁骨
- 迷齿——牙釉质深入到牙齿质深层。
- 孔:内鼻孔——犁骨和犁骨的外侧。下颞孔——颌肌穿行,翼骨、外翼骨和方轭骨、方骨之间。
- 肺鱼
- 软骨化。只保留副蝶骨、翼骨、犁骨等少数骨骼。
- 翼骨发达。和前关节骨组成磨齿。
- 原始辐鳍鱼
- 方骨。
- 上翼骨和蝶骨区有可动关节。
- 舌颌骨。续骨。
- 副蝶骨
- 【真骨鱼类】腹肌室,副蝶骨和脑匣底部分离。
- 前犁骨【真骨鱼类和鲟鱼】位于中央,单个,为两块骨合并
- 腭方软骨——后端腭骨,前端“翼骨”【内翼骨、外翼骨、后翼骨】
- 古总鳍鱼
- 下颌
- 总鳍鱼——齿骨、上隅骨、隅骨、夹板骨、前关节骨、喙状骨
- 高等硬骨鱼——齿骨、关节骨、隅骨
- 软骨脑颅
-
两栖类:
-
软骨脑颅
-
古两栖类
- 枕骨区:基枕骨,单枕髁。
- 耳骨区——前耳骨、后耳骨,卵圆窗接受耳柱骨 【来自于舌颌骨】 传来的震动。
- 蝶骨区:底墙形成基蝶骨,和两侧翼骨连结。侧墙不骨化
- 脑匣前端:蝶筛骨【鱼类左右眶蝶骨愈合】,腹部有眶间隔。
-
现代两栖类
- 枕骨区:外枕骨,双枕髁。
- 耳骨区——前耳骨。无后耳骨的种类被 外枕骨 代替或愈合。
- 蝶骨区:没有基蝶骨。有尾类和无足类有眶蝶骨;无尾目没有眶蝶骨,有蝶筛骨。
-
古两栖类 现代两栖类 基枕骨、单枕髁 外枕骨、双枕髁 前耳骨、后耳骨,卵圆窗接受耳柱骨传来的震动 只具有前耳骨。没有后耳骨的种类被 外枕骨 代替或愈合。 底墙基蝶骨,和两侧翼骨连接、侧墙不骨化 没有基蝶骨。有尾类和无足类有眶蝶骨;无尾目没有眶蝶骨,有蝶筛骨。 脑匣前端 有蝶筛骨,腹部有眶间隔。
-
-
膜颅顶罩
- 古两栖类
- 吻骨消失、鼻骨和腭骨加长,额骨之后是顶骨和后顶骨。
- 围眶骨——同总鳍鱼类。
- 颊部——鳞骨,边缘有方轭骨
- 上颌缘——前颌骨、颌骨、轭骨、方轭骨。方轭骨构成上颌缘和围眶骨,和总鳍鱼【不参与上颌缘】不同。
- 外鼻孔缘——前颌骨、颌骨、泪骨、鼻骨
- 耳裂——内缘为间颞骨、上颞骨、案骨;外缘为鳞骨。古两栖类头骨的标志性特征。
- 颞骨下有空隙——颌肌附着。向前通向眼眶,向后通向后颞孔。
- 现存两栖类:顶罩膜骨大量缺失。
- 有尾类——轭骨和方轭骨消失,只留下前颌骨、颌骨、额骨、顶骨、前额骨、鼻骨、鳞骨
- 无尾目——边缘完整,前颌骨、轭骨、方轭骨;围眶骨消失。额骨和顶骨愈合成为额顶骨。顶罩背侧只留下鼻骨、额顶骨、鳞骨。
- 无足目:完整。
- 古两栖类
-
腭罩
- 古两栖类
- 排列同总鳍鱼
- 副蝶骨显著扩大,到基蝶骨腹面。
- 颚部和脑匣愈合
- 腭方软骨后方骨化为方骨、中部形成上翼骨 和蝶骨区连结。
- 下颞孔加大,为颌肌通路
- 口盖——成对的犁骨、腭骨、翼骨、外翼骨和副蝶骨
- 内鼻孔周缘——前颌骨、颌骨、犁骨、腭骨
- 具有外翼骨——原始性。
- 现存两栖类:骨化不佳,外翼骨消失,翼骨缩小不再和犁骨相接。
- 有尾类:副蝶骨宽阔,腭骨消失,犁骨大
- 无尾目:
- 副蝶骨十字形、腭骨退化为棒状,翼骨和犁骨缩小。
- 翼间空洞——副蝶骨和翼骨之间,可容纳眼球和软腭。
- 无足目完整
- 古两栖类
-
下颌
- 古两栖类:同古总鳍鱼
- 现存两栖类:
- 有尾类:每侧三块膜骨,外侧齿骨,内侧前关节骨和夹板骨
- 无尾目:每侧一块齿骨和一块 隅-夹板骨。
-
-
爬行类
- 软骨脑颅
- 枕骨区——四块骨。单一的枕髁。【哺乳类方向进化的,基枕骨退化】
- 蝶骨区——
- 前端出现蝶筛骨。
- 眶蝶骨不骨化,并形成膜质眶间隔,脊底型。
- 侧蝶骨【耳前小柱形成。蛇、鳄鱼、鸟类】
- 脑匣和腭罩愈合【龟、鳄鱼】——基蝶骨的基翼突消失,枕骨区和方骨联合等
- 膜颅顶罩
- 丢失了间颞骨
- 丢失了上颞骨、案骨、后顶骨【杯龙类外】
- 耳裂消失,方骨扩大延伸来支持鼓膜前端。
- 颞孔出现。
- 无孔类
- 下孔类:哺乳类,爬行类
- 上孔类:鱼龙类、蛇颈龙类
- 双孔类:鳞龙类、初龙类【蜥蜴下方颞孔后墙和下墙膜骨消失。蛇类连上墙也消失并连结眼窝】
- 顶骨孔【喙头蜥、一些蜥蜴】
- 腭罩:
- 杯龙类:犁骨、腭骨、翼骨、外翼骨、副蝶骨
- 蜥蜴和蛇类:方骨周围膜骨消失,可以自由活动。某些蛇类有杠杆式关节——方骨、翼骨、外翼骨、腭骨、颌骨
- 龟鳖类:腭罩外翼骨消失,脑匣底墙和腭罩愈合,方骨粗大并和枕骨愈合。
- 鳄鱼:方骨不可动。次生腭(水中捕食)——前颌骨、颌骨、腭骨、翼骨。和哺乳类(口腔咀嚼)——不含翼骨区别。
- 下颌:
- 原始爬行类:
- 外侧——齿骨、隅骨、上隅骨
- 内侧——夹板骨、喙状骨、前关节骨。
- 初龙类:隅骨之间有孔洞。
- 哺乳类方向:齿骨增大。
- 原始爬行类:
- 软骨脑颅
-
鸟类:
- 共同:
- 脊底型
- 发达眶间隔
- 侧蝶骨
- 单个枕髁,基枕骨。
- 不同:
- 脑发达——侧壁变为底墙,膜颅顶罩参与脑匣背部侧墙。膜颅顶部圆拱形,脑颅腔大。
- 头骨薄而轻
- 鸟喙——前颌骨、颌骨、鼻骨。
- 方骨可以活动。
- 假鳄类——双颞孔消失;眶前孔留有痕迹;牙齿退化消失;下颌骨穿洞在许多种类消失;巩膜骨发达。
- 腭罩:缩小变形不完整。犁骨愈合为单个犁骨棒;副蝶骨吻突。翼骨和腭骨、基蝶骨、方骨连结。分为古腭型、新腭型。
- 共同:
-
哺乳类:
- 软骨脑颅:
- 枕骨区——愈合。两个枕髁。枕骨大孔移到腹下方。
- 耳骨区——前耳骨、后耳骨愈合为为耳骨。耳囊移动到头骨的腹侧。隅骨演化为鼓泡,为鼓膜附着。听骨。
- 蝶骨区——基蝶骨。基蝶骨和前蝶骨、眶蝶骨、翼蝶骨【为腭方软骨的上翼骨骨化愈合形成,和原始硬骨鱼类没有同源关系】愈合成为蝶骨。
- 筛骨区——中筛骨【灵长类、食肉目、啮齿目;前蝶骨前方;筛板和侧方的眶蝶骨形成】,鼻甲骨。
- 膜颅顶罩:
- 兽孔类:
- 下颞孔
- 颧弓——轭骨、鳞骨
- 哺乳类:
- 眼眶和颞孔的后墙消失【后眶骨】,两室相通。外鼻孔愈合。
- 消失——前额骨、后额骨、眶后古、上颞骨、方轭骨。
- 愈合——后枕骨愈合为枕骨。鳞骨、耳骨和鼓泡愈合为颞骨。
- 兽孔类:
- 腭罩:
- 副蝶骨消失
- 翼骨缩小,和蝶骨愈合,成为蝶骨小翼
- 犁骨合并为鼻间隔下部。
- 腭骨发达。次生腭——前颌骨、颌骨、腭骨。使得口腔咀嚼和呼吸分开。
- 方骨——砧骨。
- 下颌:
- 单一齿骨构成。
- 关节骨——锤骨。耳柱骨——镫骨。
- 多样性
- 单孔类
- 没有泪骨
- 轭骨消失
- 有前额骨和后额骨
- 围耳骨不发达。有鼓骨环。
- 有袋类:
- 鼓泡没有或由翼蝶骨构成,而不是鼓室骨。
- 有胎盘类:
- 低等种类眼眶和颞孔相连。高等种类再次出现隔墙【灵长类完全】
- 单孔类
- 软骨脑颅:
-
内脏弓的比较解剖
- 文昌鱼:咽弓不分节。
- 圆口类:
- 软骨构成
- 鳃笼不分节
- 舌软骨
- 原始无颌类:9对鳃弓,8对鳃裂
- 鱼类:上下颌出现。前两对鳃弓被改造和破坏。唇软骨和吻突?
- 四足动物:
- 舌颌骨——听骨。哺乳类方骨和关节骨——听骨。
- 舌器——舌骨体,舌骨角【人类中和耳囊区愈合,茎突】。
- 喉头软骨
肌肉系统
- 组织学上,分为骨骼肌、平滑肌、心肌。运动学上,起点,止点,原动肌,对抗肌。屈,伸、内收、外展、旋前、旋后、提、降、收缩、开大。
- 骨骼肌的胚胎发生:简要地说,体节肌——中胚层——体节——生肌节。鳃肌——中胚层——侧板——侧板中胚层。眼球外在肌——耳前体节。鳃下肌——耳后体节【枕颈体节】。具体地说,体节肌原本在背侧,迁移到腹侧,被一条结缔组织——白线隔开。成肌细胞融合——多核的肌细胞。鳃肌因为有咽囊突出导致分节【鳃节肌】,当然,这并不能和体节——生肌节产生的骨骼肌分节情况对应。
- 骨骼肌的分类:简要地说,分为中轴肌和附肢肌。具体地说,中轴肌 包括头部肌肉、躯干和尾,膈肌。附肢肌 包括鳃节肌,皮肌。
- 鳃肌就是鳃节肌,但不是鳃下肌。
肌肉系统的演化——分为体节肌,鳃节肌和皮肌。
体节肌
体节肌包括中轴肌和附肢肌。附肢肌来自于肌节芽。
中轴肌——躯干肌、尾肌、头肌、膈肌
- 【出现】鳃下肌——在圆口类出现。侧隔——是在软骨鱼出现的【横突延申】,与之对应,出现了轴上肌和轴下肌的分化,并且出现了背肋【水平生骨隔】来源。羊膜类侧隔消失。斜肌前身——软骨鱼类侧纵束在腹鳍前方连接,纤维向腹前方延伸。肋间肌前身——硬骨鱼类轴下肌可以附着到腹肋上。躯背肌出现分化——无尾两栖类分化为横突间肌、椎弓间肌。背最长肌——无尾两栖类。轴下肌分化——有尾两栖类。无尾两栖类减少,爬行类再增加。这和肋骨的运动有关。肋间肌——爬行类。横突棘肌系——爬行类。髂肋肌系——哺乳类。
- 圆口类出现了 鳃下肌。肌节和肌隔都连接到脊柱和腹膜,深入真皮。肌节直立。
- 软骨鱼类出现了折叠的肌节,出现了侧隔,肌隔向内连接到脊柱和肋骨。出现了轴上肌和轴下肌的分化。轴上肌分为背纵束;多数情况下不明显分化,称为躯背肌。轴下肌分为侧纵束和腹纵束。有些板鳃类具有腹直肌。
- 从四足动物开始,轴下肌,附肢肌逐渐发达。
- 有尾两栖类成体具有轴下肌分化——浅外斜肌、深外斜肌、内斜肌、横肌。轴上肌不分化。腹直肌是斜肌演化来的。
- 无尾两栖类轴上肌分化为横突间肌、椎弓间肌。出现了背最长肌。但是轴下肌层数减少。这和肋骨退化相适应。
- 爬行类开始缺少侧隔。爬行类轴上肌出现了横突——棘肌系。背最长肌分裂。轴下肌类似有尾两栖类,胸部出现了肋间肌。斜肌分化。
- 鸟类轴上肌弱。轴下肌腹部没有横肌,斜肌也缩减。
- 哺乳类轴上肌略有缩减,出现了髂肋肌系【髂棘肌分为髂肋肌、背最长肌、背棘肌】。轴下肌在腹部特化。腹直肌衍生出许多肌肉。
- 鱼类尾肌发达,且轴上肌和轴下肌同等发达。
- 四足动物的尾肌的轴下肌被腰带隔断。因此轴上肌比轴下肌发达。有尾两栖类和爬行类尾肌基部膨大,但这是后肢肌的一部分。鸟类和蛙类没有尾,但是有残存的尾肌。哺乳类尾肌减少。陆地脊椎动物尾肌常常发育为肛门括约肌。
- 眼球肌 由 耳前体节 发育而来。包括下颌前体节、下颌体节、舌体节。分别由脑神经III,IV,VI支配。
- 下颌前体节 分化为上直肌、下直肌、内直肌、下斜肌。下颌体节 分化为上斜肌。舌体节 分化为外直肌。大多数的脊椎动物都如此。例外是七鳃鳗的下直肌被脑神经VI支配而不是III。
- 四足动物的上睑提肌同上直肌来自于 下颌前体节,被脑神经III支配。眼球缩肌【某些蛙类和爬行类】和瞬膜运动肌【鸟类】来自于 舌体节,被脑神经IV支配。
- 鳃下肌来自于 耳后体节。在羊膜动物中被脑神经XII支配,在非羊膜动物中被脑神经和脊神经共同支配。包括颈直肌。颈直肌衍生出舌骨胸骨肌、甲状舌骨肌、胸骨甲状肌、頦舌骨肌、肩甲舌骨肌。
- 哺乳类的舌肌并不直接来自于鳃下肌的向前延伸,而是重新从间充质开始合成的。
- 膈肌只出现在哺乳类。上面有动静脉和食管的穿孔。肌肉一般只分布在边缘,中央膜质。
附肢肌
- 板鳃鱼类的附肢肌来自于肌芽。但是高等脊椎动物的附肢肌来自于肢芽,难以确定是否来自于肌节。
- 板鳃鱼类的附肢肌来自于肌芽。每个肌节先分为两个初级肌芽,然后再分为四个次生肌芽。
消化系统
- 消化管的基本结构包括粘膜、粘膜下层、肌层、浆膜。粘膜又包括上皮层、固有层、粘膜肌层
- 舌头上有粘膜形成的舌乳头。包括丝状乳头、菌状乳头、轮廓乳头、叶状乳头。兔子多叶状乳头,人类多前三种。
- 牙齿的结构:釉质、齿质、齿骨质。牙齿通过齿骨质和齿周韧带和颌骨连接。齿质由无数的齿小管和基质组成。
- 食管前半段是横纹肌,后半段是平滑肌。但是反刍动物的食管全都是横纹肌。
- 哺乳动物胃的壁细胞分泌盐酸,主细胞分泌胃蛋白酶原。但是在低等动物,这两种职能由同一种细胞担任。胃可以分为胃底、胃体、胃小弯、胃大弯、贲门、幽门。
- 小肠绒毛的中心是固有层。大部分乳糜被中央乳糜管吸收。
- 人的唾液腺有腮腺、下颌下腺、舌下腺。其中腮腺是浆液性腺体,后二者是混合性腺体。
- 人的肝脏被胃十二指肠肝韧带联系。被冠状韧带联系。肝小叶是肝脏的结构功能单元。肝小叶内有肝细胞,组成单列的肝板。胆汁流入胆小管,小叶间胆管、总胆管。
- 消化系统的发生。肝脏和胰脏是原肠伸出的盲囊形成的。要区分口道和口腔,它们被口板隔开。内鼻孔脊椎动物口腔最前,包括鼻囊和垂体囊。同样的有肛道。文昌鱼的口和脊椎动物的口没有同源关系。
- 牙齿的发生。颌骨产生唇沟,向前产生唇,向后产生齿龈。齿龈产生齿板,釉器。釉器是牙齿产生的原基,产生釉质。齿板上常具有齿栅,用来换齿。
- 表皮齿见于圆口类、无尾两栖类幼体、原兽亚纲。
- 咽囊的分化。哺乳动物咽囊分化情况:I鼓室——II腭扁桃体、颈动脉体——III甲状旁腺和胸腺——IV甲状旁腺——V后鳃体。
- 胃肠的分化。需要转动。背系膜随之转动产生网膜囊,最后产生大网膜,在小肠和大肠的腹面。此后胃肠扭转。产生肠袢、盲肠突。在盲肠突产生盲肠和阑尾。胚胎时期都是泄殖腔,因为产生了尿直肠隔,分开产生肛门和泄殖窦。
- 肝脏前端通过冠状韧带连接到横膈膜,通过镰刀韧带连接到腹壁。文昌鱼的肝盲囊可能是肝脏的雏形,但是它既没有肝脏的功能,也没有纤毛腺体上皮。在胰脏的发生过程中,会产生背胰和腹胰。后来腹胰通过扭转到背部和背胰融合。
- 口裂——有颌类才具有真正的口裂。哺乳类的唇和颊发达。
- 腭部——总鳍鱼、肺鱼、两栖类出现了内鼻孔。爬行类开始不再平坦,出现了突起;并且出现腭褶,内鼻孔后移。哺乳类出现横棱。
- 舌——圆口类的舌和其他脊椎动物没有同源关系。鱼类是基舌骨突出覆以粘膜形成的。多鳍鱼和肺鱼出现肌肉质的舌。四足动物的舌是鳃下肌分化的。有的鸟类具有舌下囊储存食物。鱼类和两栖类的舌被第一对脊神经支配。其后则被脑神经VII和V支配。
- 口腔腺——两栖类出现唾液腺【颌间腺、鼻间腺】。鸟类具有角腺。雨燕的唾液腺发达,可以筑巢。哺乳类具有 腮腺、下颌下腺、舌下腺。偶蹄类还具有臼齿腺,犬科还具有眶腺。
- 牙齿——银鲛和肺鱼牙齿为齿板状,为取食贝类的适应。辐鳍鱼类为锥形。古总鳍鱼类和两栖类具有迷齿。蛙类上颌有齿,蟾蜍没有齿。蜥蜴、蛇类、喙头蜥具有腭齿。槽生齿【鳄鱼】、侧生齿【蛇类、大多数蜥蜴】、端生齿【少部分蜥蜴和喙头蜥】。
- 低等脊椎动物的牙齿上有闪光质,和哺乳动物的釉质并不同源。闪光质来自于中胚层,而釉质来源于外胚层。
- 反刍类不具有上颌门齿。三尖齿是哺乳动物牙齿形式的演化基础,食虫。食肉类具有裂齿。食草类牙齿方化,高冠齿。
- 鸟类食管下段常形成膨大的嗉囊和喉囊。
- 文昌鱼、圆口类、银鲛、肺鱼和一些真骨鱼类不具有胃。鱼类和两栖类的胃J形。羊膜类出现了胃大弯和胃小弯。鸟类分化为腺胃和肌胃。反刍类的皱胃是真正的胃。
- 螺旋瓣是盾皮鱼、某些早期硬骨鱼类、七鳃鳗、板鳃类具有的。大多数硬骨鱼类和低等辐鳍鱼类具有幽门盲囊。
- 羊膜动物出现盲肠。鸟类具有一对盲肠。哺乳类一般一个盲肠。
- 软骨鱼类、两栖类、爬行类、鸟类、单孔类具有泄殖腔。
- 鲤科肝脏分散。七鳃鳗成体不具有胆囊。鸟类和哺乳类有的种类不具有胆囊——鸽子、奇蹄目、鲸鱼、偶蹄类。
- 板鳃类胰脏独立于消化管之外。辐鳍鱼类分散。哺乳类一般具有两条胰管。
排泄系统
-
前肾的命运:
- 在 软骨鱼和有尾两栖类,前肾管纵分为 吴氏管 和 牟勒氏管。
- 其他动物,前肾管变成吴氏管。而 牟勒氏管 由腹膜内陷包卷形成。
-
膀胱的类型:
导管膀胱 泄殖腔膀胱 尿囊膀胱 无膀胱 硬鳞鱼、硬骨鱼 肺鱼、两栖类、单孔类。 龟鳖类、喙齿蜥、哺乳类、部分蜥蜴。 圆口类、软骨鱼、蛇类、鳄鱼、部分蜥蜴、大多鸟类 -
文昌鱼的肾管来自外胚层。
-
硬骨鱼具有头肾。输精管和输卵管是各自生殖腺壁延伸而成的,和排泄系统没有关系。
-
软骨鱼雄性肾前端变为莱氏腺,后端尾肾发出副肾管。中肾管输精,而副肾管输尿。
-
有尾两栖类肾前端变为附睾肾。
-
羊膜类雌性吴氏管变为卵巢冠纵管,中肾变成卵巢冠和卵巢旁体。雄性中肾变为旁睾和附睾附件。
神经系统
- 灰质——神经元胞体、树突、神经胶质细胞、几乎没有髓鞘的轴突。皮质、核团都是灰质。基底神经节或者纹状体是包埋在白质中的灰质。
- PNS具有神经节,也是神经细胞胞体集中的地方。脑神经、脊神经的神经节,是感觉神经元所在的地方;植物性神经的神经节则是换元的地方。
- 网状结构是灰质和白质交织的地方。种系发生上比较古老。
- 无脊椎神经系统包括网状神经系统、梯状神经系统、链状神经系统,在节肢动物中甚至具有脑化。半索动物背神经索前伸进入领和吻。尾索动物成体只具有一个神经节。头索动物神经系统比较发达,但不具有脑泡。
- 脊椎动物神经系统的个体发生。神经板、神经褶、神经嵴、神经管。神经管形成CNS。神经嵴形成PNS的神经节。神经管形成后,管壁向成神经细胞和成胶质细胞分化。
- 脊髓的发生。管壁分化为室管膜上皮、套层【组成脊髓的灰质】、边缘层【没有细胞核】。成神经细胞产生多极神经元。
- 脑的发生。通过腹曲、脑峡的分区,产生前脑、中脑、菱脑。后来嗅觉、视觉、听觉分别和前脑、中脑、后脑联系,从而导致脑向背部扩展产生了大脑半球、顶盖、小脑。前脑分化为端脑、间脑。端脑有嗅球、大脑半球;间脑有脑上体【松果器】,产生视泡、视神经【所以从某种角度上说,视神经是“中枢”神经系统的一部分】,并由脑漏斗产生垂体。
- 总的来说,前脑分为端脑和间脑。端脑——大脑半球【皮层、髓质】、基底神经节、大脑皮层、嗅脑【嗅叶、嗅束、嗅球、梨状叶、海马】。间脑——丘脑上部、丘脑【视丘】、丘脑下部【灰结节、漏斗、脑下垂体、视交叉、乳头体】、视神经、松果体。中脑——四叠体、顶盖、视叶、被盖、大脑脚。菱脑分为后脑和末脑。后脑——哺乳动物脑桥、小脑【小脑半球、蚓部、绒球】。末脑——延髓。
- 脑室。侧脑室、第三脑室、中脑水管【在这里向每一侧产生中脑腔。】、【低等动物在前部具有后脑腔】第四脑室、末脑腔。
- 脉络丛——室管膜构成了脉络丛上皮。脉络丛是 间脑前段和延髓顶部 只保留室管膜上皮层,并和软膜融合产生的。
- 脑脊液循环。脑脊液在左右侧脑室脉络丛产生。室间孔、第三脑室,中脑导水管、第四脑室。在第四脑室流出进入蛛网膜下腔,通入静脉窦,颈内静脉。
- 脊髓有颈膨大和腰膨大。高等脊椎动物有腹裂、正中沟、背隔、连合。脊椎动物脊髓长度逐渐缩短,代表头化。脊髓有背角、腹角、侧角。灰质的排列在高等脊椎动物中大多为H形。背角大多是联合神经元,腹角大多是运动神经元。
- 脊髓灰质分区有感觉柱、运动柱。并有分躯体和内脏。内脏柱位于中部,躯体柱在内脏内侧。感觉柱位于背面,运动柱位于腹面。白质叫做索。有外侧索、背索、腹索。腹索还可以分为束。
- 真正管状的脊髓出现在圆口类。鱼类出现背裂,出现成对的灰质腹角。两栖类出现腹裂,灰质开始出现H形。鸟类缺少终丝。腰膨大背面分离,具有菱形窦,容纳大的胶质细胞。哺乳动物出现背外侧沟和腹外侧沟。
总
- 颌关节的类型。分为舌接型、自接型、颅接型。
- 初生腭、次生腭和初生颌、次生颌。初生颌只在软骨鱼类出现,麦氏软骨和腭方软骨;在其后的类群骨化称为次生颌。初生腭由。次生腭由前颌骨、上颌骨、腭骨【鳄鱼的次生腭还有翼骨参与,适应水下生活。哺乳动物的次生腭没有翼骨参与,适应口腔咀嚼。】组成。
- 颞弓、颧弓、颞窝的组成。颞弓是由。
- 无脊椎动物的进化关系。
- 合胞体上皮:轮虫、线虫、腹毛、棘头。
- 骨质鳞片的进化。
- 真皮属于致密结缔组织而不是疏松结缔组织。
- 分泌方式。包括局部分泌、顶浆分泌【汗腺、乳腺】、全浆分泌【皮脂腺】。
- 表壳虫-几丁质。有孔虫-钙质。鳞壳虫-硅质。
- 脊椎凹型。双凹型见于鱼类、鲵类、壁虎。前凹型 两栖类。后凹型 爬行类。异凹型见于鸟类颈椎。双平型见于哺乳动物。
- 三个听小骨和韦氏小骨。三个听小骨分别是锤骨【关节骨】、砧骨【方骨】、镫骨【舌颌骨】。韦氏小骨见于鲤鱼,是最前方的三个脊椎骨变来的,连接鳔和内耳。
- 弧胸型和固胸型。本质的区别是前乌喙骨是否交叉。弧胸型见于蟾蜍和雨蛙。固胸型见于其他蛙类。
- 附肢骨模式图。
- 哺乳动物的三种足型。
- 鸟类附肢骨的特化。
- 软体动物的肌痕。闭壳肌粗大,缩足肌前后都具有,只有前方具有前伸足肌。
- 鱼的枕髁。硬骨鱼枕髁。软骨鱼枕髁。
- 无椎体的原始类型。
- 参差型椎体。青蛙的第七个脊椎第六个躯椎是前凹形,荐椎双凸型,第八个脊椎是双凹型。蛙科、树蛙科、姬蛙科具有。蟾蜍全为前凹型。
- 哺乳动物C7的例外。海牛C6。树獭C9。
- 眼球肌。
- 肋间外肌和肋间内肌。肋间外肌走向腹后方,起点在上方肋骨靠中间的部分,终点在下方肋骨靠边缘的部分,吸气。肋间内肌走向腹前方,呼气。
- 鸟类的肌肉系统。适应飞翔——胸大肌用来把翅膀往下动,胸小肌用来把翅膀上抬。
- 平胸总目的代表物种。
- 呼吸色素的总结。
- 动脉系统的不对称发生于爬行类。
- 婴儿的循环系统。肺不起到呼吸作用,而是胎盘起到呼吸作用。心房之间具有卵圆孔。肺动脉和体动脉之间具有动脉导管。具有静脉导管。
- 一些特殊循环。
- 葡糖、肌酐和肾清除率。
- 奇静脉和半奇静脉。奇静脉位于右侧,半奇静脉位于左侧。他们都是后主动脉在哺乳动物的残留。
- 扩大消化系统面积的方式。
- 骆驼反刍胃的特化——不具有瓣胃。瘤胃、网胃可以存水。
- 燕雀类唾液腺具有消化酶。
- 马、兔消化主要依赖盲肠。牛的消化主要依靠反刍胃。
- 哺乳动物的进化关系。
- 有尾两栖类动脉的特化。第5动脉弓保留并入体动脉弓。体动脉和肺动脉之间具有动脉导管。部分有尾两栖类颈动脉和体动脉之间有颈动脉导管。
- 虾的解剖。
- 哺乳动物皮肤的一般结构。
- 哺乳动物锁骨的情况。
动物生理学
细胞信号转导
- 胆碱能通路、胺能通路、氨基酸能通路。
- 胆碱能通路 包括 脑神经运动核、脊髓前角支配随意运动;脑干网状结构非特异性上行激动系统;脊髓后角、脊髓侧丘脑、大脑皮层特异性感觉投射;交感神经节前,副交感神经节前和节后神经元支配内脏运动。
- 胺能通路:交感神经节后,脑桥,延髓。
- 氨基酸能通路。在脑中广泛分布。兴奋性的有Glu,Asp。抑制性的有GABA,Gly,牛磺酸。
- MAchR的分类
- M2,M4:Gi。心脏窦房结、房室结、心室肌、心房肌;收缩力下降【弱心】
- M1,M3,M5:Gq。胃肠道、输尿管的平滑肌和汗腺;收缩力增加;血管平滑肌舒张(促进 NO 合成,非直接)。【其他】
- 肾上腺素能受体:
- α1:Gq.平滑肌收缩;主要是血管平滑肌,存在于皮肤、胃肠道、肾动脉、立毛肌;细支气管、膀胱逼尿肌(比β2的松弛作用小)
- α2:Gi.小肠平滑肌舒张。
- β:Gs.
- β1:心脏组织,心肌正变力、正变时、正变传导。【强心】
- β2:胃肠道、支气管、膀胱平滑肌松弛,骨骼肌血管舒张。【其他】
- β3:脂肪组织脂肪分解、膀胱逼尿肌松弛。【脂肪】
- 不同类型受体对 E 和 NE 的敏感性:
- α1:NE。α2:=。β1:NE。β2:E。β3:=。
- NE:α > β1 > β2。通过β1增强心脏的收缩力,加快心率;通过α1受体让血管收缩。
- E:α = β1 = β2。通过β1增强心脏的收缩力,加快心率;通过 β2 舒张骨骼肌和肝脏血管。
消化吸收和能量代谢
-
消化管的管壁包括 黏膜、黏膜下层、肌层、外膜。黏膜包括 上皮、固有层、黏膜肌层。口腔食管和肛门的上皮是复层鳞状上皮,胃、小肠大肠的上皮是单层柱状上皮。
-
肠神经系统包括黏膜下神经丛和肌间神经丛。交感神经节后纤维主要和肠神经系统联系而不是直接和平滑肌联系。迷走神经支配横结肠和以上的消化道,盆神经支配降结肠和以下的消化道。
-
消化道是体内最大最复杂的内分泌器官。G细胞分泌胃泌素【延缓胃排空】。I细胞分泌缩胆囊素【分泌酶】。S细胞分泌促胰液素【分泌水和HCO-】。K细胞分泌抑胃肽【刺激胰岛素】。
G细胞 I细胞 S细胞 K细胞 胃泌素 缩胆囊素 促胰液素 抑胃肽 延缓胃排空 分泌酶 分泌水和HCO- 刺激胰岛素 -
慢波起源于Cajal细胞。位于环形肌和纵行肌交界处。
-
吞咽中枢 位于延髓和脑桥下段的网状结构。呕吐中枢 位于延髓。排便中枢 位于脊髓腰骶段。
吞咽中枢 呕吐中枢 排便中枢 延髓和脑桥下段的网状结构 延髓 脊髓腰骶段 -
胃的 容受性舒张 是迷走-迷走反射调节的,节后纤维释放的递质不是Ach而是 VIP【血管活性肠肽】。
-
胃排空速率 糖类>蛋白质>脂质。
-
肠胃反射 抑制胃的运动,被酸、脂肪、脂肪酸、高渗溶液刺激。
-
小肠的 分节运动 以环形肌为主。上部频率高,下部频率低。消化间期小肠有 移行性复合运动,起源于胃的中部。
-
副交感神经 在小肠运动中的作用没有胃运动中重要。胃泌素、P物质、5-HT可以 加强 小肠运动。促胰液素、生长抑素、VIP可以 抑制 小肠运动。
加强小肠运动 抑制 胃泌素、P物质、5-HT 促胰液素、SST,VIP -
消化腺。胃腺、小肠腺、大肠腺都是黏膜上的上皮细胞组成的。唾液腺有三对——腮腺【舌咽神经,下泌涎核】、下颌下腺、舌下腺【面神经、上泌涎核】。胃 中 外分泌 有贲门腺、泌酸腺【主细胞、壁细胞、粘液细胞】、幽门腺。内分泌 有G细胞、D细胞、ECL细胞【分泌组胺,可以刺激胃排空】、EC细胞【5HT】。粘液的主要成分是糖蛋白。内因子由壁细胞分泌。小肠腺包括 伯氏腺 和 李氏腺,李氏腺分泌酶【唯一分泌 肠激酶】。大肠腺 不分泌酶只分泌粘液。
-
刺激胃液分泌 的物质有Ach,胃泌素、组胺。生长抑素、前列腺素、上皮生长因子可以 抑制 胃液分泌。头期 胃蛋白酶含量高,肠期 胃液分泌量最大。
刺激胃液分泌 抑制 Ach,胃泌素、组胺 生长抑素、前列腺素、上皮生长因子 -
鸽子、大象、鹿、马、驴子、大鼠等生物没有胆囊。它们的总胆管括约肌阻力很小,肝分泌的胆汁直接通过总胆管进入十二指肠。
-
十二指肠和空肠上端 吸收糖 的能力最强。单糖吸收速率葡萄糖、半乳糖>果糖>甘露糖。经过SGLT1进入肠上皮,经过GLLUT2入血。部分单糖通过水解转运进入细胞。
-
新生哺乳动物少量蛋白质可以通过胞吞作用进入细胞。二肽或三肽转运速率通常超过AA。经过H+-肽同向转运体。氨基酸有的通过Na+梯度转运。
-
脂质消化产物吸收的限速步骤是和胆盐形成混合微胶粒。CM通过淋巴进入血液。十二指肠和空肠吸收脂质的能力最强。
-
大部分的 水 在小肠下段被吸收。Na+ 主要在空肠被吸收。Ca++主要在十二指肠被吸收,涉及钙结合蛋白。Fe++在十二指肠和空肠被吸收,吸收Fe++,需要把食物中的Fe+++通过VC 还原。和转铁蛋白结合胞吞入胞。
水 Na+ Ca++ Fe++、脂质、糖 小肠下段 空肠 十二指肠 十二指肠和空肠 -
蛋白质的特殊动力效应最强。热天少吃蛋白质。
渗透调节和排泄
-
鸟类和哺乳动物的肾利用逆流倍增产生高渗尿。爬行动物不能产生高渗尿。在没有水的情况,骆驼不排尿而是把尿素储存在组织中。
-
肾也是重要的内分泌器官。它可以产生 肾素、钙三醇、促红细胞生成素EPO。
-
肾小囊有两层上皮细胞。内层是后肾小管盲端折返内陷形成的。肾小囊壁层和脏层具有 肾囊腔,和肾小管的管腔相通。
-
近端小管上皮锥体形,髓袢上皮扁平,远端小管上皮立方,集合管上皮立方或柱状。
-
皮层肾单位和髓旁肾单位。
皮质肾单位 髓旁肾单位 入球小动脉粗 = 出球小动脉离开肾小囊后第二次分成毛细血管网 出球小动脉离开肾小球后,分成网形小血管和直小血管。 -
近球小体包括 近球细胞【分泌肾素】、致密斑【感受远端小管的Na+浓度】、球外系膜细胞【吞噬和收缩】。
-
大部分肾血流在肾皮质部。肾血流量的 自身调节 只 涉及肾皮质的血流量。
-
肾小球滤过率 是单位时间内肾产生的超滤液 量。用菊糖或者肌酐测定。
-
滤过膜包括 毛细血管内皮细胞、基膜【水合凝胶,对肾小球滤过率起到决定作用】、肾小囊脏层足细胞。分子量70000。电荷性质:带负电的透性小,中性次之,正电的透性大【滤过膜上有唾液蛋白,带有负电。】
-
有效滤过压 = 肾小球毛细血管压-血浆胶体渗透压【向静脉内大量灌注生理盐水,胶体渗透压减小】-肾小囊内压。
-
急性肾小球炎 减少了有效滤过膜面积,少尿水肿;但同时增大了肾滤过膜的通透性,导致血尿和蛋白尿。肾血流量主要通过影响 滤过平衡。
-
肾小管对水的重吸收:近端小管 吸收了 67% 的水。
-
-
肾小管对 Na+ 和 Cl-的重吸收:
- 近端小管前半段:吸收70%的Na+。
- 近端小管后半段:吸收10%的Na+。Cl-顺浓度梯度【因为前半段HCO3-的吸收远大于Cl-吸收】,Na+顺电位梯度。通过细胞旁路和跨上皮。涉及H+和甲酸盐F-的再循环。交换体。HF入胞。
- 髓袢降支细段:Na+扩散入胞。
- 髓袢升支粗段:NKCC同向转运体【Na+,2Cl-进入组织。K+进入小管腔,1个Na+顺电位差通过细胞旁路进入组织】。总的来说,2Cl-主动进入重吸收,1Na+主动重吸收,1Na+被动重吸收。对水不通透。
- 远曲小管起始段:Na+主动重吸收。
- 远端小管后端和集合管:主细胞和闰细胞。主细胞吸收Na+【顺电位进入细胞】和水,分泌K+。闰细胞分泌H+。
-
-
肾小管对HCO3-的重吸收:不易透过管腔膜。通过CO2的形式进入小管。主要在近端小管吸收。由于 近端小管 的Na+-H+交换,HCO3-的吸收 远大于 Cl-吸收。
-
K+的重吸收:最后排出的K+基本是远端小管分泌出来的。
-
葡萄糖重吸收的部位仅限于近端小管。尿素重吸收主要在近端小管。代谢产物均在近端小管被主动分泌。蛋白质通过胞吞进入细胞。
-
H+分泌和Na+-H+交换:分泌H+,对应的HCO3-进入血液,Na+顺电位差扩散进入细胞。
-
K+分泌和Na+-K+交换:只有当Na+的主动重吸收存在时,才能分泌K+。K+的分泌是顺电位差的。
-
NH3生成主要在近端小管。和H+结合产生NH4+。
神经感觉
- 适应 是 外周 感受器的一种调节,而 习惯化 是 中枢 内突触效率的修饰导致的。
- 脊髓丘脑前束——粗触觉和压觉:
- 脊神经后根节【上升或下降若干个脊髓节段,故切断脊髓不能使得相应的皮节感觉全部丧失】
- 脊髓灰质的后角内。在脊髓 交叉 到对侧。
- 丘脑。
- 大脑皮层的躯体感觉区。
- 脊髓丘脑侧束——痛觉和温度觉。最后投射到中央后回的中上部和旁中央小叶后部。
- 三叉神经束——头面部浅感觉。
- 三叉神经半月神经节
- 周围突——感觉支;
- 中枢突——三叉神经感觉根,进入脑桥。
- 升支,传导 触觉,三叉神经 主核
- 降支,传导 痛觉 和 温度觉,三叉神经 脊束核
- 交叉,三叉丘系,在内侧丘系的背侧
- 三叉神经半月神经节
- 意识性深感觉传导通路:
- 脊神经节,假单极神经元
- 周围突
- 中枢突:升支、降支、侧支
- 薄束——躯体下部——部分延髓薄束核、楔束——躯体上部——部分延髓楔束核
- 内侧丘系,交叉
- 腹后外侧核,丘脑皮层束。
- 大脑皮层中央后回中上部,旁中央小叶后部和中央前回。
- 脊神经节,假单极神经元
- 头面部意识性深感觉:三叉神经中脑核或三叉神经节——丘脑——中央后回。
- 非意识性深感觉
- 脊神经节。中枢突——脊髓中间内侧核或背侧核。
- 脊髓小脑前束、后束。
- 丘脑是大脑皮层兴奋的最直接来源。
- 丘脑核团:
- 感觉接替核群
- 腹后内侧核——三叉丘系——中央后回
- 腹后外侧核——内侧丘系,脊髓丘脑束——中央后回
- 腹外侧核——小脑红核丘脑束——中央前回
- 内侧膝状体——外侧丘系听觉——颞叶
- 外侧膝状体——视神经——枕叶
- 联络核群:背内侧核、后外侧核、丘脑枕
- 非特异性投射核群:髓板内核群(中央中核)。
- 感觉接替核群
- 简化眼和曲率半径:节点 n 到瞳孔的距离即 曲率半径。并非眼球前后径而是从物像到节点的位置。曲率半径增大看远物。
- 视杆细胞和视锥细胞都没有产生动作电位的能力。以 电紧张 的形式扩布。
- 内淋巴含有 高浓度的K+ 和 低浓度的Na+,由血管纹分泌,具有耳蜗内电位。
神经运动
-
运动神经元被称为运动系统的最后公路。
-
脑干的下行运动通路:
-
内侧下行通路:发生古老,终止于 腹内侧部。和躯体姿势控制有关。
- 前庭脊髓束——前庭迷路关于平衡和姿势反射
- 网状脊髓束——维持姿势
- 顶盖脊髓束——头部和眼球运动
前庭脊髓束 网状脊髓束 顶盖脊髓束 前庭迷路关于平衡和姿势反射 维持姿势 头部和眼球运动 -
外侧下行通路:终止于 背外侧部。和四肢特别是手的定向运动有关。
- 红核脊髓束——指挥手和手指的运动。【但人类的红核脊髓束并不发达,功能被皮层脊髓束取代】
-
-
大脑运动皮层包括 初级运动皮层,【前运动皮层、辅助运动皮层——前运动区,复杂运动的协调和计划】。
-
大脑皮层的下行运动通路:
- 皮层脊髓束——躯体肌肉和四肢肌肉。
- 皮层脊髓 侧束——下降纵贯脊髓全长,和前脚外侧部联系。四肢远端肌肉。和技巧有关。
- 皮层脊髓 前束——只下降到胸部,通过中间神经元和前脚内侧部联系。四肢近端肌肉和躯干肌肉。古老。
- 皮层脑干束——面部肌肉
- 此外还有皮层网状脊髓束,皮层红核脊髓束等。
- 非哺乳类脊椎动物基本上没有皮层脊髓束和皮层脑干束。
- 不全型麻痹——运动传导通路受损。对应肌肉的肌张力下降。
- 皮层脊髓束——躯体肌肉和四肢肌肉。
-
小脑和基底神经节。小脑 通过对皮层运动指令和实际运动情况对比反馈。基底神经节 接受 所有 皮层区域的传入,传出到运动计划的额叶。
-
锥体系和锥体外系。
- 锥体系——皮层脊髓束和皮层脑干束。发动随意运动。
- 锥体外系——经典锥体外系、旁锥体系【锥体系发出分支进入皮层下中枢】、皮层起源锥体外系【皮层通过皮层下中枢接替】。调节肌紧张,调整姿势,协调肌群运动。
-
梭内肌纤维的 中间部分 没有收缩能力。梭内肌纤维可以分为 核袋纤维【Ia,板状末梢】和 核链纤维【II和Ia,蔓状末梢】。感觉纤维有Ia和II。γ梭内肌纤维的末梢有蔓状末梢和板状末梢。
-
高尔基腱器官被 Ib 传入纤维支配,连在和骨骼肌相连的胶原纤维的细束上。
-
肌梭感受外力牵拉。腱器官感受主动收缩。
-
行走要依赖局部脊髓环路——中枢模式发生器。此外脊髓以上的中脑行走区等也参与了行走控制。
-
运动计划,运动编程和运动执行。
- 运动计划:大脑皮层联络区、基底神经节、小脑外侧。
- 运动编程:大脑初级运动皮层、小脑
- 运动执行:脑干、脊髓。
-
前运动区——前运动皮层和辅助运动皮层。失用症——不能进行复杂的运动。
- 辅助运动皮层——双侧性。近端直接,远端间接【摧毁初级运动皮层再刺激辅助运动皮层,远端肌肉没有收缩】。
- 前运动皮层——躯体中轴肌肉和四肢近端肌肉。
-
小脑可以分为前庭小脑、脊髓小脑、皮层小脑。
-
前庭小脑 接受 平衡觉和视觉 传入——躯体平衡和眼球运动功能紊乱,眼球震颤。
-
脊髓小脑——通过顶核控制内侧下行系统,控制躯体中轴和肢体近侧的肌肉活动。通过间位核控制外侧下行系统,控制肢体远端的肌肉活动。共济失调【随意运动不准确】,意向性震颤,肌张力减退。
-
皮层小脑——不接受输入。通过齿状核经过腹外侧核到皮层运动区和前运动区。参与随意运动的计划和编程。远侧肢体的肌张力下降,运动起始延缓。
-
前庭-眼反射——运动学习。
-
前庭小脑 脊髓小脑 皮层小脑 平衡觉和视觉传入 控制肌肉活动 随意运动的计划和编程 躯体平衡和眼球运动功能紊乱,眼球震颤 共济失调、意向性震颤、肌张力减退 远侧肢体的肌张力下降、运动起始延缓
-
-
基底神经节。鸟类以下的动物,基底神经节 是运动的最高中枢。基底神经节包括 尾核、壳核【新纹状体,输入】、苍白球【旧纹状体,内侧部输出】、底丘脑核、黑质。
-
基底神经节 不接受 外周感受器,而是接受大脑皮层和黑质的传入【黑质纹体束,可以增强直接通路,抑制间接通路】。
-
基底神经节的传出通过腹前核和腹外侧核。包括直接通路和间接通路。
- 直接通路:从尾核、壳核直接到苍白球内侧。抑制腹前核和腹外侧核。易化大脑发出运动。
- 间接通路:从尾核、壳核先到苍白球外侧,再到苍白球内侧。去抑制。抑制大脑发出运动。
-
帕金森病:肌紧张 过强而运动过少。静止性震颤。黑质 投射到纹状体的 多巴胺能神经元 退行性病变导致。
-
亨廷顿病:肌紧张过少而 运动 过多。尾核和壳核 的 GABA和胆碱能神经元 退行性病变导致。
-
去大脑僵直:α-僵直,前庭脊髓束。γ-僵直,网状脊髓束。
-
自主神经系统 节前B纤维,节后C纤维。
-
中枢对内脏活动的调节:
- 脊髓:血管张力,发汗,排尿,排便,勃起。
- 延髓:心血管运动,呼吸,咳嗽,喷嚏,吞咽,唾液分泌,呕吐。
- 脑桥:呼吸调整中枢、角膜反射中枢
- 中脑:瞳孔对光反射中枢,视听探究反射中枢
脊髓 延髓 脑桥 中脑 血管张力,发汗,排尿,排便,勃起 心血管运动,呼吸,咳嗽,喷嚏,吞咽,唾液分泌,呕吐 呼吸调整中枢、角膜反射中枢 瞳孔对光反射中枢,试听探究反射中枢 -
下丘脑:体温——视前区-下丘脑前部。摄食中枢——外侧部。饱中枢——腹内侧核。饮水中枢——外侧区。内分泌——视上核和室旁核。昼夜周期——视交叉上核。防御反应区——腹内侧核。逃避反应——背侧区。
-
边缘系统:大脑半球内侧面皮层和脑干连结部和胼胝体旁的环周结构——边缘叶。
内分泌
-
儿茶酚胺类舒张支气管和收缩血管需要糖皮质激素的 允许作用。
-
垂体前叶来自于中胚层,和下丘脑没有直接的神经纤维联系,通过垂体门脉毛细血管联系。垂体后叶则可以视为下丘脑的延伸。
-
嗜酸性细胞 分泌 GH 和 PRL。嫌色细胞 分泌阿黑皮素原【包括 ACTH, MSH, LPH, END】。嗜碱性细胞 产生 TSH, LH, FSH。
嗜酸性细胞 嫌色细胞 嗜碱性细胞 GH,PRL ACTH,MSH,LPH,END TSH,LH,FSH -
GH 还可以通过 肝脏产生 生长介素【一种 IGF-I】 执行功能。巨人症、肢端肥大症、侏儒症。胃饥饿素 可以也可以促进 GH 分泌,导致摄食行为的发生。
-
雌激素对 PRL 有 拮抗作用。高浓度 PRL 导致 闭经泌乳综合征。
-
GH,PRL,ACTH是垂体的 三大应激激素。
-
ACTH, 糖皮质激素的分泌具有 节律性——凌晨多,午夜少。褪黑素的分泌同样具有 昼夜节律性,晚上多,白天少。
-
在女性,FSH促进卵泡的生长、发育,雌激素分泌。LH促进排卵和黄体形成。在男性,FSH促进生精细胞的成熟,LH促进间质细胞分泌睾酮。
-
OT 和 VP 结构类似。在鱼类、两栖类、一些鸟类合为加压催产素。OT是射乳反射的重要激素。VP缺失导致尿崩症。酒精 抑制VP分泌。
-
甲状腺的神经主要调节血流量,和内分泌功能调节关系不大。
-
尽管甲状腺激素为胺类激素,但其合成来自于甲状腺球蛋白的水解。被储存在细胞外的滤泡腔。生理剂量的甲状腺激素促进蛋白质合成,但过多则导致蛋白质水解,如甲亢。呆小症。大脖子病。对心血管系统有 甲状腺危象——心动过速和心肌过度收缩导致的心力衰竭。
-
降钙素降低血磷降低血钙。甲状旁腺增加血钙降低血磷。1,25-VD-(OH)2增加血钙增加血磷。1,25-VD-(OH)2的产生必须经过VD先后在肝脏和肾脏的两次羟基化。
-
胰岛素通过 RTK——P13K——PIP2——PKB发挥作用。促进 GLUT4。胰岛素缺乏导致酮血症和酸中毒,渗透性利尿。NO也可以促进胰岛素分泌。胰高血糖素促进,SST抑制;促胰液素、胃泌素、缩胆囊素、抑胃肽都可以促进。
-
高浓度的胰高血糖素可以强心。交感神经和迷走神经都促进胰高血糖素的分泌。
-
肾上腺皮质球状带分泌盐皮质激素;束状带分泌糖皮质激素;网状带分泌性激素。
-
应急反应和应激反应:交感-肾上腺髓质介导应急反应。肾上腺的皮质和髓质都参与应激反应。
-
肾素——血管紧张素——盐皮质激素系统。
生殖
- 睾丸实质包括 曲细精管【生成精子】和 间质细胞【合成并分泌雄激素】。曲细精管包括 精原细胞 和 支持细胞。注意区分支持细胞和间质细胞。
- 支持细胞对生精的作用:支持保护营养;形成血睾屏障;分泌雄激素结合蛋白、金属结合蛋白、维生素结合蛋白;吞噬。支持细胞还可以分泌抑制素;表达芳香化酶生成雌激素。还可以通过胞质运动将逐渐分化的精子推入管腔。
- 哺乳动物精子形成的 变形期 中,鱼精蛋白代替了组蛋白和DNA紧密结合【令人感叹,鱼类没有鱼精蛋白】;形成顶体【高尔基体先形成泡状结构,然后逐渐增大,凹陷为双层帽状覆盖在核的头端,称为顶体】;鞭毛、轴丝、线粒体鞘;细胞质大量丢失。
- LH脉冲式释放,FSH稳定释放。LH刺激间质细胞生成睾酮,睾酮直接反馈抑制LH。FSH刺激支持细胞生精,支持细胞分泌 抑制素 间接反馈抑制FSH。
- 卵巢的内分泌作用。滤泡细胞和颗粒细胞分泌雌激素。双重细胞学说【LH刺激下在滤泡细胞产生雄激素,FSH刺激下在颗粒细胞再生成雌激素】。排卵后,黄体细胞分泌孕酮和较多的雌激素。
- 雌激素可以增多HDL,减少LDL;促进 钠水潴留。孕激素可以提高基础体温,抑制排卵、促进钠水潴留。
- 若 持续给予GnRH,将会导致受体下调从而抑制性激素分泌。
- 月经周期的内分泌调控。
- 卵泡期早期。FSH,LH分泌——雌激素增多——反馈抑制FSH——只有一个优势卵泡继续发育。
- 月经周期中期。雌激素增多——反馈促进FSH,LH,GnRH——LH峰出现并排卵。
- 黄体期。雌激素分泌下降——LH作用下黄体发育——孕激素、雌激素增多——LH,FSH减少。
生态学
群落的组成和结构
- 生物群落的定义——在相同时间聚集在同一地段上的各物种种群的集合。先强调时间,后强调空间
- 地植物学就是植物群落学。
- 群落交错区具有边缘效应——种的数目和一些种的密度增大。
- 机体论和个体论。机体论把群落比作是一个有机体,看成是一个自然单位,封闭群落。Clements。个体论学派认为群落是不存在明显边界的,开放群落。Gleason,Ramensky,Whittaker。
- 最小面积——基本上能够表现出某种群落类型植物种类的最小面积。
- 优势种——有明显控制作用。建群种——优势层的优势种。亚优势种。伴生种。偶见种。关键种——有重要的和不相称的作用,虽然并不占优势,这是和优势种的区别,比如传粉昆虫。
- 群落的数量特征。多度——个体数目的多少。密度——样地内某种植物个体数占据总植物个体数的百分比。盖度,分盖度,总盖度,盖度比——某种植物占最大盖度物种的盖度的百分比,基盖度,显著度——乔木的基盖度。频度——出现样方数占总样方数量的百分比。Raunkiaer频度定律——A>B>C>=D<E。群落的均匀度和A和E的比值大小有关。重要值=相对密度+相对频度+相对基盖度。
- 辛普森多样性【\(D=1-\sum P_i^2\)】和香农威那多样性【\(H=-\sum P_i \log P_i\)】,香农威那多样性单位是nit,bit,dit。
- alpha多样性——群落中,beta多样性——群落间,gama多样性——最广阔的地理尺度。
- 解释物种多样性空间关系的学说。进化时间学说、生态时间学说、空间异质性学说、气候稳定学说、竞争学说、捕食学说、生产力学说。
- 关联系数的计算【\(V\frac{ad-bc}{\sqrt{(a+b)(c+d)(a+c)(b+d)}}\)】。
- 生活型——体态相似、适应特点也相似。潮湿热带——高位芽。沙漠、草原——一年生植物。温带、北极——地面芽植物。
- 多数浮游动物其实趋向弱光。在白天它们在水的深层,在晚上它们上升到水的表面。
- 同资源种团是等价种。海洋生物种间竞争比陆地生物多,大型生物比小型生物多。植食性昆虫种间竞争少。
- 干扰和抽彩式竞争【各物种竞争能力相同,但是先占据断层的物种相比后占据断层的物种占据优势】。高多样性取决于对断层的抽彩式竞争。
- 演替和多样性——多样性开始较低,演替中途增加,到达顶极多样性又稍微降低——中度干扰假说,藻类的多样性取决于波浪干扰下石头移动的频率而不是石头的大小。
- 小演替和抽彩式竞争。小演替有多个建群种,抽彩式竞争只有一个建群种。
- 鸟类的多样性相比于植物种数,不如和取食高度多样性关系更紧密——对于鸟类来说,植被的分层结构比物种组成更加重要。
- 海岛的物种数-面积曲线【\(S=CA^Z\)】。Macauthur的平衡说——平衡时迁入率和灭绝率相同【综合考虑岛屿大小和陆地远近。灭绝率不被远近影响】。
- 资源越多;平均生态位宽度越小;平均生态位重叠越大;饱和度越大——物种数越多。
群落的动态
- 偏途性波动——是气候和水分条件的长期偏离引起的若干个优势种明显变更的结果。
- 任何一个群落形成,必须要经过植物的传播、植物的定居和植物的竞争。
- 沙丘是湖水退却后逐渐暴露出来的。因此沙丘上的基质是原生裸地。
- 根据P/R比,可以区别自养性群落和异养性群落。
- 演替过程——促进模型、抑制模型和耐受模型。适应对策演替理论、资源比率理论、等级演替理论。
- 演替顶极——单元顶极论【只有气候顶极】、多元顶极论【如土壤顶极、地形顶极】、顶极——格局假说【连续的顶极类型。优势顶极——分布最广泛并且位于格局中心的顶极】。
群落的分类和排序
- 分类——机体论,封闭群落。排序——个体论,开放群落。
- 群丛【基本单位。各层片结构和优势种相同。】、群系【中级单位。建群种相同】、植被型【建群种生活型相同】。英美学派优势种原则,群系最大。法瑞学派考虑植物区系的亲缘关系。北欧学派以基群丛作为基本单位。前苏联学派以群丛、群系、植被型为主要单位。
- 按照属性排列实体叫做正分析【Q】。按照实体排列属性叫做逆分析【R】。利用环境因素排序叫做直接排序,利用植物群落本身属性排序叫做间接排序。间接排序主要适用主分量分析法。
发育生物学
神经管的形成和图示化
- 外胚层的主要衍生物:
- 表面外胚层【高水平的骨形态发生蛋白】——表皮、毛发、指甲、皮脂腺、嗅上皮、口上皮【包括垂体前叶、牙釉质、颊上皮】、晶状体、角膜
- 神经嵴【中等水平】——周围神经系统【包括施万细胞、神经胶质细胞、交感神经系统、副交感神经系统】、肾上腺髓质、黑素细胞、面部软骨、牙本质
- 神经板/神经管【低水平】——脑、神经垂体、脊髓、运动神经元、视网膜。

 浙公网安备 33010602011771号
浙公网安备 33010602011771号